i když těkavé sloučeniny s agonistickou aktivitu byly testovány proti ORs, konkrétní chemické látky se silným agonistický účinek na komáry, 2-(4-Ethyl-5-(pyridin-3-yl)-4H-1,2,4-triazol-3-ylthio)-N-(4-ethylphenyl)acetamid (VUAA1), otevřel oblasti výzkumu (Jones et al., 2011; Taylor et al., 2012). Pozdější studie Taylor et al., (2012) poskytl důkazy o chemických látkách odvozených od VUAA1, jako je VUAA4, schopných zvýšit svůj agonistický účinek 10krát na ORco od A.gambiae, h. virescens a harpegnathos saltator. Zajímavé je, že autoři uvedli, že jakákoli změna na amidových substituentech způsobí úplnou ztrátu agonistické aktivity. To poskytuje užitečné informace o strukturálních požadavcích agonistů a vztahu struktury a aktivity mezi analogy VUAA a ORs. Konečně, navzdory zvýšené agonistické aktivitě chemikálií VUAA, jeho relativně vysoká molekulová hmotnost (367, 47 g mol−1 pro VUAA1) vs., Volatilní agonisté, jako je bombykol (238,42 g mol−1), činí přímé Volatilní doručení VUAA něco, co není proveditelné. S tím na mysli, hledání menších strukturálních analogů představuje zajímavé zaměření výzkumu.
Antagonismus Na ORco Podjednotky
Spolu se studiem VUAA související s analogy, které mohou působit jako silné agonisty, zablokování ORco tím, že antagonisté se také objevila na průvodce semiochemicals a pesticidů design., Proto, strukturální analog VUAA1, VU0183254 (2-(4-Ethyl-5-furan-2-yl-4-triazol-3-ylsulfanyl)-1-fenothiazin-10-yl-ethanone), bylo hlásil, že inhibují ORco reakci, působí jako alosterický modulátor u A. gambiae a narušuje uznání agonistů, jako je např. eugenol v komplexu OR65/ORco (Jones et al., 2012). Jiné vuaa-strukturální analogy byly také hlášeny jako antagonisté. Příkladem je N-,2-substituované triazolothioacetamide sloučeniny OLC3 a OLC12, že narušuje ORco odpověď v podobné módě v C. quinquefasciatus, A. gambiae, D. melanogaster a O., nubilalis, což naznačuje konzervované vazebné místo v ORco (Chen a Luetje, 2012). S ohledem na inhibici ORco jako slibná strategie narušit chování hmyzu, zdá se, že následné úsilí by se mělo zaměřit na sloučeniny s nižší molekulovou hmotností, než VUAA-odvozené antagonisté. Například, OX1a (232 g mol−1), tryptamin (160.22 g mol−1) a isopropyl-cinnamát (190.24 g mol−1) byly hlášeny antagonistickým účinkem na ORco (Chen a Luetje, 2013, 2014; Tsitoura et al., 2015) se zhruba poloviční nebo menší molekulovou hmotností než VUAA1., Nicméně budoucí použití těchto antagonistů by mělo být pečlivě studováno, protože zablokování konzervovaného ORco může ovlivnit nejen škodlivý hmyz, ale i prospěšné.
Kromě toho, antagonisty efekt zkoumali in vitro, důkazy na behaviorální úrovni podporuje myšlenku, že strukturální analogy feromony mohou fungovat jako antagonisté. Například Sellanes et al., (2010) uvádí, inhibice sexuální reakce v medovice můra Cryptoblabes gnidiella, když strukturální analogy, (Z)-9-tetradecenyl formiátu a (Z)-11-hexadecenyl mravenčanu, byly přidány na syntetický sexuální feromon, (Z)-11-hexadecenal a (Z)-13-octadecenal, v aerodynamickém tunelu testy. Tento feromonový antagonistický účinek byl později potvrzen v terénních testech, kde se zachycení samců C.gnidiella snížilo v závislosti na dávce. Feromonový antagonismus byl také hlášen pro B.mandarinu, předchůdce B. mori (Daimon et al., 2012)., Jejich zjištění potvrdit bombykol jako sexuální feromon, a bombykal a bombykyl acetát jako antagonisté, který silně inhibuje přitažlivost mužů v poli na sex feromon bombykol. Nedávno byly hlášeny důkazy o feromonovém antagonismu pro čenichovou můru Herpetogramma submarginale. Když (Z)-13-hexadecenol byl přidán do jeho pohlaví feromon, (Z) – 13-hexadecenylacetát, významně snížil počet mužů zachycených v poli (Yan et al., 2015)., Feromonový antagonismus se zdá být založen na rozdílech v chemické funkční skupině, jako jsou alkoholy, aldehydy a estery v závislosti na druhu hmyzu. Nicméně, antagonisty vliv těchto strukturních analogů nemusí být kvůli ORco inhibici ale specifičnost ORx na antagonisty. Nedávná studie naznačuje, že OR16 helicoverpa armigera je schopen specificky rozpoznat feromonový antagonista (z) – 11-hexadecenol (Chang H.et al., 2017). Autoři podpořili konkrétní roli OR16 s ohledem na to, že H., armigera samice vypouštějí antagonisty sloučenina spolu s jeho sexuální feromon ((Z)-11-hexadecenal a (Z)-9-hexadecenal) jako strategie, aby se zabránilo non-optimální krytí s nezralými muži. Výjimečně, když OR16 byl sražen genomu editační techniky CRISPR/Cas9 a H. armigera mužů byly testovány elektrofyziologické a behaviorální testy, ne EAG odpověď byla zaznamenána a muži se snažili spářit s nezralé samice.
receptory odorantu vs., Vazebné Proteiny: Klady a Zápory pro Hmyzí Škůdce Řízení
Pro případ OBPs, ligand specifičnost a mechanismy OBPs představují kontroverzní aspekty, což se zdá být silně závislé na metodách používaných pro měření ligand afinitu. Například bylo hlášeno, že PBP, jako jsou ty z molů P.xylostella a eogystia hippophaecolus, mohou vázat jak složky feromonu, tak analogy (Sun et al., 2013a; Hu et al., 2018). To naznačuje, že následní hráči, jako je ORs, by mohli zvýšit specifičnost a citlivost příjmu odorantu., Nedávné důkazy podporují, že koexprese PBPs a PRs může zvýšit citlivost vůči feromonům. Například několik kombinací z PR1 – 4 a PBP1-4 bylo použito k testování jejich reakce na sexuální feromonové složky můry Chilo supressalis (Chang et al., 2015). Autoři zjistili významné zvýšení citlivosti reakce na (Z)-11-hexadecenal, když byly PR4 a PR6 spolu exprimovány s PBP4., Ačkoli interakce těchto proteinů by mohla vzniknout nová úroveň výzkumu jako pest control cíle, různé párování PRs a Pbp vrhnout světlo na složitosti čichového systému u hmyzu, takže přístup obtížný úkol pro velkou skupinu sloučenin a proteinů test. Navzdory výše uvedenému jsou hmyzí OBPs malé molekulární velikosti se snadnou produkcí rekombinantních proteinů, což z nich činí oblíbené cíle strukturálních studií a rychlého vazebného screeningu., Například, ligand screening s OBPs by mohly umožnit identifikaci chemické vlastnosti pro lepší vázání, jako je délka řetězce, molekulární objem, funkční skupiny, a bond nenasycenosti. Tyto, v kombinaci s novými proteinové struktury, predikce, metody používané v designu léčiv a protilátek, jako je homologie modelování, simulace dynamiky, a molekulární dokování, může místo bodnutí OBPs v oblíbené pozici nejvzdálenější regiony jako cíle pro vývoj řídících agentů v ochraně proti škůdcům.
hmyzí Ori se zdají být specifičtěji naladěni na odoranty než OBPs., Vyšší specificita projevená ORs a šance na aktivaci / inhibici specifických receptorů pro dané chování činí tyto proteiny jako atraktivní cíle pro manipulaci s chováním škůdců. Proveditelnost inhibice komplexu ORx/ORco nebo ORco antagonisty zahrnuje slibnou strategii k narušení chování specifického pro hmyz, jako je Páření pomocí pohlavních feromonových receptorů. Nedostatek strukturálních informací je však překážkou při používání hmyzu ORs jako cílů pro předpovědi semiochemické aktivity., Tabulky 1, 2 shrnují počet OBPs a ORs, které byly u druhů hmyzu identifikovány transkriptomem(tj. Většina dosud studovaných hmyzu má podle genomových studií alespoň dvakrát ORs než OBPs. Kromě toho dochází k rozsáhlému rozšíření ORs v sociálním hmyzu z řádu Hymenopteranů, jako je medonosná včela a.mellifera se 170 ORs (Weinstock et al., 2006) a mravenci Solenopsis invicta a Cerapachys biroi s 400 A 506 ORs (Wurm et al., 2011; Oxley et al., 2014)., Podobně je rozšíření nebo patrné také u některých zemědělských škůdců, jako je brouk červené mouky T. castaneum s 265 ORs ve srovnání s 47 OBPs (Richards et al., 2008). To je náročný úkol pro cílové NEBO výběr spolu s obtížnosti pro funkční expresi transmembránových proteinů, jako jsou nejvzdálenější regiony, aby se na obrazovce velké množství ligandů. Aproximace důležitých vlastností v obou OBPs a ORs jsou shrnuty v tabulce 3.
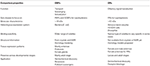
Tabulka 3., Přibližné srovnání hmyzu ORs a OBPs podle vlastností.
Další Perspektivy
funkční charakteristika hmyzu Nejvzdálenějších regionů, jakož i jejich prokázané role v čichu hmyzu mají vrhnout světlo na citlivost a specifičnost těchto hmyzu-specifické proteiny. Tyto pokroky dále zvýší jejich proveditelnost jako cíle kontroly škůdců pochopením mechanismů molekulárního rozpoznávání a kombinačních interakcí s OBPs., Na druhé straně bude pokračovat současné masivní úsilí v identifikaci a závazné charakterizaci OBPs u několika zemědělských důležitých druhů hmyzu a poskytne více informací o jejich funkcích ve fyziologii hmyzu. Tak, tato recenze navrhuje jako hlavní výhodu pro OBPs přes nejvzdálenější regiony, dostupnost 3D crystal a NMR struktury, které s navazující přístupy, jako je homologie modelování (v případě potřeby), molekulární dokování a molekulová dynamika, by upřesnit hledání bioaktivních látek., To poslední v doplnění s ligand afinitu měření urychlí studium hmyzu OBPs být přehodnocena jako cíle pro semiochemical discovery a nástroje pro návrh super-ligandy v pest control management.
vzhled a vývoj insekticid rezistence u hmyzích škůdců vedly k intenzivnímu výzkumu na hmyz čichu a mechanismy, které se podílejí na nervové procesy. Je dobře prokázáno, že řada receptorů a enzymů v CNS hmyzu je cílem vývoje rezistence vůči insekticidům (Obrázek 1)., Bylo prokázáno, že acetylcholinesterázy (AChE) v rozpustné podobě zajišťuje odolnost vůči organické sloučeniny fosforu a karbamátu insekticidy, působí jako bioscavengers (Lee et al., 2015). Podobně, více insekticid mechanismy rezistence byla prokázána u mšice M. persicae, zahrnující carboxylesterases, sodíkové kanály, γ-aminomáselné kyseliny (GABA) a nAChR (Bass et al., 2014)., Jako důležité komponenty v hmyzu periferii nervového systému a klíčoví hráči v chování hmyzu, a to jak hmyzu OBPs a nejvzdálenější regiony představují alternativní cíle pro identifikaci sloučenin s semiochemical činnosti (nebo agonistický účinek) a nástroje pro návrh silní antagonisté pro zvýšení požadované behaviorální reakce hmyzích škůdců a snížit používání insekticidů a následné odpor.
autorské příspěvky
HV napsaly sekce o struktuře OBP a ORs, vyvinuté tabulky a obrázek., J-JZ pojal myšlenku na recenzní článek, napsal hlavní sekci, jako je úvod, funkce OBPs a struktura, jakož i srovnání OR-OBP.
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
poděkování
autoři by rádi poděkovali FONDECYT 3170433., J-JZ je vděčný za finanční podporu ze strany Northeast Normal University a Jilin University, Čína za jeho volno ke studiu v Číně. Rothamsted Research obdrží grant s přidaným financováním z biotechnologie a biologických věd (BBSRC), Spojené království.
Gonzalez, F., Witzgall, P., and Walker, W. B. (2017). Antennální transkriptomy tří tortricidních můr odhalují domnělé konzervované chemosenzorické receptory pro sociální a habitatové čichové podněty. Věda. Rep. 7, 1-12. doi: 10.,1038 / srep41829
PubMed Abstract / CrossRef Full Text / Google Scholar
Kaissling, K. E. (2013). Kinetika čichových reakcí může do značné míry záviset na interakci odorantu a receptoru a na deaktivaci odorantu postulované pro detektory toku. J. Comp. Fyziol. Neuroethol. Smyslové Nervové Chování. Fyziol. 199, 879–196. doi: 10.1007 / s00359-013-0812-z
PubMed Abstract / CrossRef Full Text / Google Scholar