Návrh a konstrukce randomizované knihovny pro prubířství Cas9 PAM předvolby
PAM knihovny obsahující randomizované sekvencí DNA okamžitě po proudu sekvence DNA komplementární k distanční vodící RNA byly generovány a použity k empiricky určit PAM uznání Typu II Cas9 endonucleases (Obr. 1)., Při fixaci cílové sekvence distanční RNA slouží randomizované báze jako substrát pro přímé odečítání specifičnosti Cas9 endonukleázy PAM. Randomizované sekvencí byly zavedeny do plasmidu DNA vektoru v PAM regionu protospacer cílové sekvence prokazující perfektní homologie s guide RNA distanční T1 (CGCUAAAGAGGAAGAGGACA). Dvě knihovny zvyšující se ve velikosti a složitosti z pěti randomizovaných párů bází (1,024 potenciál PAM kombinace) do sedmi randomizovaných párů bází (16,384 potenciál PAM kombinace) byly generovány., Randomizace knihovny 5 bp byla zavedena syntézou jediného oligonukleotidu obsahujícího pět náhodných zbytků. Single-stranded oligonukleotid byl přeměněn na double-stranded šablony pomocí PCR (Další soubor 1: Obrázek S1A), klonován do plasmidu vektor (Další soubor 1: Obrázek S1B) a transformovány do E. coli jak je popsáno v Metodách části., K zajištění optimální náhodnost v 7 bp PAM knihovny, velikosti a složitosti knihovna byla snížena syntézou čtyř oligonukleotidů každá obsahuje šest náhodných reziduí plus sedmý pevné zbytky obsahující G, C, A, nebo T, resp. Každý ze čtyř oligonukleotidů byly jednotlivě převedeny do dvouvláknové DNA, klonovaných do vektoru pTZ57R/T, jak je popsáno v Metodách oddíl a transformovány do E. coli jak je popsáno pro 5 bp knihovna., Po transformaci, plasmidová DNA byla získána zpět, a v kombinaci z každé ze čtyř 6 bp PAM knihovny generovat randomizované 7 bp PAM knihovna obsahující 16,384 možné PAM kombinace. Pro obě knihovny, začlenění náhodnost byla ověřena hluboké sekvenování; zkoumání nukleotidové složení v každé pozici PAM regionu pomocí pozice frekvence matrix (PFM) (Metody, část a ) (Další soubor 1: Obrázek S2A a B)., Distribuce a frekvence každé sekvence PAM v 5 bp a 7 BP randomizovaná knihovna PAM jsou uvedeny v dalším souboru 1: Čísla S3 a S4.
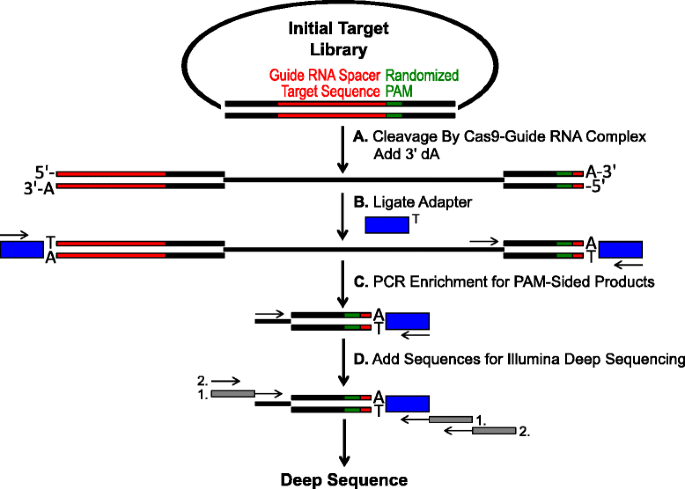
Schéma pro identifikaci PAM preference Cas9 štěpení in vitro. počáteční plasmidová knihovna s randomizovaným PAM (green box) je štěpena komplexem Cas9 a přidávají se převisy 3′ dA. B adaptéry s převisem 3 ‚ dT (modrá krabička) jsou ligovány na oba konce štěpného produktu., C primery jsou využívány k obohacení o Pam-oboustranné štěpené produkty PCR., d Po PCR obohacení, DNA fragmenty jsou čistí a Illumina kompatibilní kotvy a čárové kódy jsou ‚sledoval-na‘ přes dvě kola PCR (šedá políčka) a Illumina hluboké sekvenován,
Prubířství Cas9 PAM předvolby
randomizované PAM knihovny je popsáno v předchozí části byly podrobeny in vitro trávení s různými koncentracemi rekombinantního Cas9 protein s předinstalovaným guide RNA za účelem testu endonukleázy Cas9 PAM preference v závislosti na dávce., Po trávení s Cas9-guide RNA ribonucleoprotein (RNP) komplexy, PAM pořadí kombinace z náhodně PAM knihovny, které podporované štěpení byly zachyceny podvazování adaptéry zdarma-končí plasmidu DNA molekuly štěpí na Cas9-guide RNA komplexu (Obr. 1a a b). Podporovat efektivní ligaci a zachycení štěpí koncích, tupým koncem, double-stranded DNA snížit generované Cas9 endonucleases byl upraven tak, aby obsahovat 3′ dA převis a adaptéry byly upraveny tak, aby obsahují komplementární 3′ dT převis., Generovat dostatečné množství DNA pro sekvenování, DNA fragmenty přechovávání PAM sekvence podporuje štěpení PCR byly amplifikovány pomocí primerů v adaptér a další přímo sousedící s PAM regionu (Obr. 1c). Výsledné PCR amplified Cas9 PAM knihovny byly převedeny do ampli-seq šablon (obr. 1D) a single-číst hluboké sekvenované z adaptéru straně amplicon., S cílem zajistit přiměřené pokrytí, Cas9 PAM knihovny byly sekvenovány do hloubky nejméně pět-krát větší než rozmanitost v počáteční randomizované PAM knihovny (5,120 a 81,920 čte pro 5 a 7 bp PAM randomizované knihovny, respektive). PAM sekvence byly identifikovány z výsledné sekvence dat, pouze o výběr těch, čte obsahující dokonalou 12 nt sekvence zápas doprovodná obou stranách 5 nebo 7 nt PAM sekvence (v závislosti na randomizované PAM knihovna slouží); zachycovat jen ty PAM sekvence vyplývající z dokonalé Cas9-guide RNA cílového místa, rozpoznávání a štěpení., Pro kompenzaci inherentní zaujatosti v počátečních randomizovaných knihovnách PAM byla frekvence každé sekvence PAM normalizována na její frekvenci ve výchozí knihovně. Vzhledem k tomu, že zde popsaný test přímo zachycuje Cas9 štěpitelné Pam sekvence, bylo pravděpodobnostní modelování použito k výpočtu konsensu PAM pro každý protein Cas9. Toho bylo dosaženo vyhodnocením pravděpodobnosti nalezení každého nukleotidu (G, C, a nebo T) v každé poloze sekvence PAM nezávisle pomocí matice polohové frekvence (PFM) (sekce metod a )., Výsledné pravděpodobnosti byly pak vizualizovány jako WebLogo .
pro zkoumání sklonu k falešným pozitivům v testu bylo vynecháno přidání komplexů Cas9 RNP do kroku trávení (obr. 1a) a test byl proveden prostřednictvím kroku obohacování PCR (obr. 1c). Jak je znázorněno v dalším souboru 1: obrázek S5A, v nepřítomnosti komplexů Cas9-guide RNA nebyly zjištěny žádné amplifikační produkty. To znamená, že výskyt falešných pozitiv je nízký a významně nepřispívá k výsledkům testu.,
PAM preference Streptococcus pyogenes a Streptococcus thermophilus (CRISPR3 a CRISPR1 systémy) Cas9 bílkovin
Za účelem ověření testu, PAM preference Streptococcus pyogenes (Spy) a Streptococcus thermophilus CRISPR3 (Sth3) Cas9 proteiny, jejichž PAM sekvence požadavku bylo dříve oznámeno , byly zkoumány. In vitro výluhy byly prováděny s 1 µg (5.6 nM) 5 bp randomizované PAM knihovna na dvou koncentracích 0, 5 a 50 nM, pre-smontované Spy nebo Sth3 Cas9 bílkovin, crRNA, a tracrRNA RNP komplexy pro 1 h ve 100 µL reakční objem., Na základě jejich frekvence v 5 bp randomizované PAM knihovny, Špionážní a Sth3 Cas9 PAM sekvence (NGG a NGGNG, respektive) na konečné koncentrace 0.40 nM a 0,11 nM při trávení, resp. Členové randomizované PAM knihovna, která obsahovala PAM sekvence, které podporované štěpení byly zachyceny a identifikovány, jak je popsáno v předchozí části. Jako negativní kontrola, vyvolávací uncleaved randomizované PAM knihovna byla předmětem sekvenování a PFM analýzy vedle těchto knihovnách vystaveny Cas9 RNP komplexy., Jak je uvedeno v Další soubor 1: Obrázek S5B a C, bez pořadí preference existují v nepřítomnosti Cas9 RNP komplex trávení jako evidentní téměř dokonalé rozdělení jednotlivých nukleotidů na každé pozici PAM v PFM stůl a nedostatek informativní obsah v WebLogo pro kontrolu. To je v ostrém zúžení s obr. 2a a b, které ilustrují složení sekvencí odvozených z knihoven strávených komplexy Spy a Sth3 Cas9 RNP. Zkoumání weblogů odvozených od PFM (obr., 2a a b)také odhalují přítomnost kanonických Pam preferencí pro proteiny Spy a Sth3 Cas9 , NGG a NGGNG. Přestože jsou preference PAM hlášené pro proteiny Spy a Sth3 Cas9 pozorovány jak u 0, 5 nM, tak u 50 nM, dochází k obecnému rozšíření specificity za podmínek trávení 50 nM. To je nejvíce patrné v pozici 2 pro Spy Cas9 protein, kde se frekvence nekanonického zbytku a dramaticky zvyšuje (obr. 2a)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
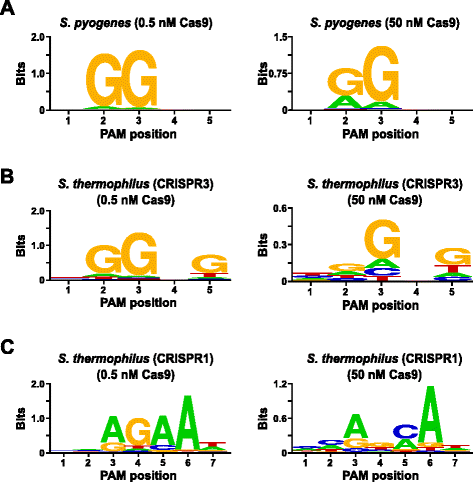
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., Frekvence nukleotidů na každé PAM pozici byl samostatně vypočítán pomocí pozice, frekvence matrix (PFM) a vyneseny jako WebLogo
Další validace assay byla provedena porovnáním PAM preferencí pro Streptococcus thermophilus CRISPR1 (Sth1) Cas9 protein, jehož PAM specifičnosti bylo hlášeno, že se rozšíří do 7 bp . Pomocí 1 µg (5.6 nM) 7 bp randomizované PAM knihovny jako šablony, Sth1 Cas9-guide RNA zažívací trakt byly provedeny na dvou koncentracích 0,5 nM a 50 nM, RNP komplexu, jak je popsáno výše., Jako ovládací prvky, Spy a sth3 Cas9 RNP komplexy byly také použity k trávení 7 bp randomizované PAM knihovny, ale pouze v jednom, 0.5 nM, RNP komplexní koncentrace. Na základě frekvence v 7 bp randomizované PAM knihovny, PAM sekvence již bylo dříve oznámeno pro Sth1 (NNAGAAW), Spy (NGG), a Sth3 (NGGNG) na konečné koncentrace 0,01 nM, 0.22 nM, a 0,05 nM., Jak je uvedeno v Další soubor 1: Obrázek S6A a B, PAM preference pro Špionážní a Sth3 Cas9 proteiny vytvořené pomocí 7 bp knihovny byly téměř totožné s těmi, které se vyrábí s 5 bp knihovny poskytují silný důkaz pro reprodukovatelnost testu. PAM preferencí pro Sth1 Cas9 bílkovin také úzce uzavřeno, že již bylo dříve oznámeno, NNAGAAW , na 0,5 nM Cas9-guide RNA komplex koncentrace (Obr. 2c)., Podobné Špionážní a Sth3 Cas9 proteiny, Sth1 Cas9 bylo schopné štěpit rozmanitější sadu PAM sekvence v reakcích obsahujících vyšší koncentrace Cas9-guide RNA komplex (50 nM), nejvýraznější byla označena ztráta G reziduí požadavek na pozici 4 a do blízkosti stejné preference pro C a bp v poloze 5 (Obr. 2c). To mělo za následek jiný konsenzus PAM než konsensus získaný při nižších koncentracích.,
posoudit, zda PAM specifičnost je nezávislý na typu guide RNA, duplexed crRNA:tracrRNA nebo sgRNA , Spy, Sth3, a Sth1 Cas9 PAM preference byly také zkoumány pomocí binární Cas9 a sgRNA RNP komplex. Trávení bylo prováděno při jediné komplexní koncentraci RNP 0,5 nM a byla provedena analýza preferencí PAM, jak je popsáno výše. Jak je uvedeno v Další soubor 1: Obrázek S7A, B, a C, PAM preference byly téměř identické, bez ohledu na typ guide RNA použit; buď crRNA:tracrRNA duplex nebo sgRNA., Kromě toho, k potvrzení, že PAM specifičnost není výrazně ovlivněno složení cílové DNA nebo distanční sekvence, sekvence na opačné straně 5 nebo 7 bp randomizované knihovna byla určena pro štěpení s různými distančními; T2-5 (UCUAGAUAGAUUACGAAUUC) pro 5 bp knihovna nebo T2-7 (CCGGCGACGUUGGGUCAACU) pro 7 bp knihovna. Spy a sth3 Cas9 proteiny předinstalované sgrna zaměřené na sekvenci T2 byly použity k výslechu 5 BP randomizovaná knihovna PAM, zatímco komplexy Sth1 Cas9-T2 sgRNA byly použity k trávení 7 BP randomizovaná knihovna PAM., Pam preference byly testovány, jak je popsáno výše. Pam preference pro všechny proteiny 3 Cas9 byly téměř identické bez ohledu na distanční a cílovou sekvenci DNA (další soubor 1: obrázek S8A, B A C).
Identifikace sgRNA a PAM preferencí pro Brevibacillus laterosporus Cas9 bílkovin
K empiricky zkoumat PAM preference pro Cas9 protein, jehož PAM nedefinované, uncharacterized Typ II-C CRISPR-Cas locus z Brevibacillus laterosporus kmen SSP360D4 (Blat) byl identifikován vyhledáním vnitřní DuPont Pioneer databází pro Cas9 orthologues., Locus (přibližně 4,5 kb) obsahoval cas9 gene schopný kódování 1,092 polypeptid, CRISPR pole obsahující sedm opakování-distanční jednotky jen po proudu od cas9 gene a tracrRNA kódování regionu se nachází proti proudu cas9 gene s částečnou homologii k CRISPR pole se opakuje (Obr. 3a). Délka opakování a rozpěrky (36 a 30 bp) je tedy podobná jiným systémům CRISPR-Cas typu II s pěti z osmi opakování obsahujících 1 nebo 2 BP mutace(obr. 3b a další soubor 1: obrázek S9)., Jiné geny obvykle nalezené v lokusu typu II CRISPR-Cas byly buď zkráceny (cas1) nebo chybějící (obr. 3a).
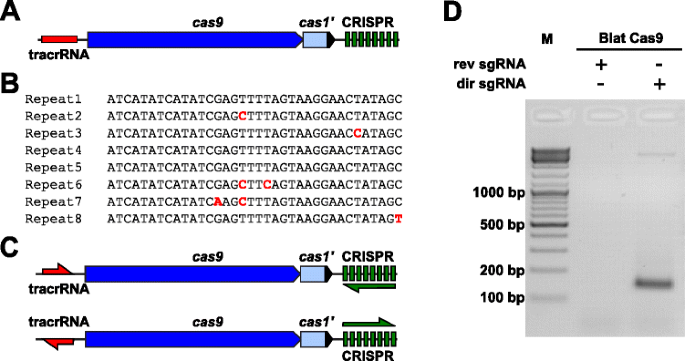
označení Typu II CRISPR-Cas prvky v Brevibacillus laterosporus SSP360D4 CRISPR-Cas systém. ilustrace oblasti genomové DNA ze systému CRISPR-Cas typu II od Brevibacillus laterosporus ssp360d4. B porovnání sekvencí opakování typu II CRISPR array identifikovaných v Brevibacillus laterosporus ssp360d4., c transkripční scénáře „direct“ a „reverse“ tracrRNA a CRISPR array pro systém CRISPR-Cas typu II od Brevibacillus laterosporus ssp360d4. d agarózového gelu s reakční produkty, což znamená, že pouze „přímé“ sgRNA (dir sgRNA), ale ne „reverzní“ sgRNA (rev sgRNA) podpora plasmidu knihovna štěpení v kombinaci s endonukleázy Cas9 pocházející z Brevibacillus laterosporus SSP360D4
guide RNA požadavek na Blat Cas9, protein byl určen generování dvou sgRNA variant., Tyto varianty byly vytvořeny tak, aby odpovídaly jak možným scénářům smyslového, tak Anti-sense exprese pole tracrRNA a CRISPR (obr. 3c) a používá sonda, která výraz scénáře podporované štěpení činnost Blat Cas9 v randomizované PAM knihovny. Jeden průvodce RNAs byly navrženy tím, že nejprve určit hranice domnělého tracrRNA molekuly na základě analýzy regionů, které jsou částečně komplementární k 22 nt 5′ konci opakování (anti-opakovat)., Dále k určení 3 ‚ konce tracrRNA byly použity možné sekundární struktury a terminátory k předpovědi oblasti ukončení v následném fragmentu. Toho bylo dosaženo screeningem na přítomnost Rho nezávislých terminačních sekvencí v DNA obklopující Anti-repeat podobný tomu popsanému v Karvelis et al. , přeměnou okolní DNA na RNA sekvenci a zkoumáním výsledných struktur pomocí UNAfold ., Výsledné sgRNAs byly navrženy tak, aby obsahovat T7 polymerázy přepis zahájení rozpoznávání signálu na 5′ konci následuje 20 nt target recognition, sekvence, 16 nt crRNA opakovat, 4 nt self-skládací vlásenka smyčka, a anti-opakujte sekvenci komplementární k opakování regionu crRNA následuje zbývající 3′ část domnělého tracrRNA. Varianta sgRNA, která obsahuje domnělý tracrna transkribovaný ve stejném směru jako gen cas9(obr. 3c) se nazývá „direct“ sgRNA, zatímco sgrna obsahující tracrRNA přepsána v opačném směru „reverzní“ sgRNA., Padesát nM Blat Cas9 sgRNA RNP komplex, pre-naložený s „přímé“ nebo „reverzní“ sgRNAs, respektive, byly inkubovány s 1 µg (5.6 nM) 7 bp randomizované PAM knihovny. Po trávení knihovny a přidání převisů 3‘ dA byly adaptéry ligovány a produkty štěpení byly zesíleny PCR (obr. 1). Analýza reakčních produktů pomocí elektroforézy agarózového gelu odhalila, že „přímá“ sgRNA, ale ne „reverzní“ sgrna podporované štěpení plasmidové knihovny (obr. 3d). Sekvence a předpokládaná sekundární struktura „přímé“ sgRNA jsou uvedeny v dalším souboru 1: obrázek S10.,
Po určení vhodné guide RNA pro Blat Cas9, PAM identifikace byla provedena obdobně jako je popsáno výše pro Spy, Sth3, a Sth1 Cas9 proteiny proti 7 bp randomizované PAM knihovna s dvou koncentracích 0, 5 a 50 nM, pre-smontované Blat Cas9 „přímé“ sgRNA RNP komplex. Jak je znázorněno na obr. 4a, PFM WebLogo PAM konsensus pro Blat Cas9 bílkovin pod 0,5 nM digest podmínek byl NNNNCND (N = G, C, A, nebo T; D = A, G nebo T) se silnou preferencí pro C, na pozici 5 PAM sekvence., Mírné preference A byl pozorován v poloze 7 a mírnou preferencí pro C nebo T na pozici 4 a G, C, nebo přes T na pozici 6 byly také zaznamenány při bližším prozkoumání PFM tabulky (Doplňující soubor 1: Obrázek S11). Podobně jako u proteinů Spy, Sth3 a Sth1 Cas9 se specificita PAM rozšiřuje, jak se zvyšuje koncentrace komplexu Cas9-sgRNA. To je nejvíce patrné v poloze 5, kde větší podíl Pam sekvencí obsahujících štěpení podpory reziduí a při 50 nM ve srovnání s podmínkami trávení 0, 5 nM.
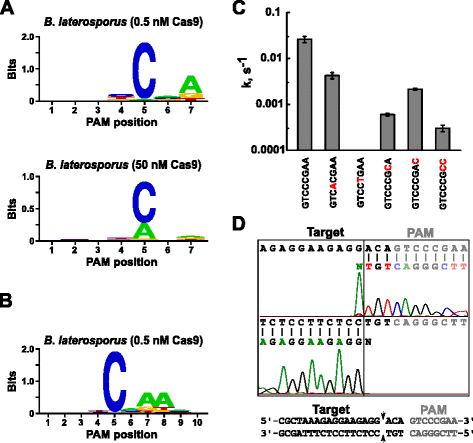
PAM preference a štěpení pozice Brevibacillus laterosporus SSP360D4 (Blat) enzym Cas9. Blat Cas9 PAM preferencí, kdy 1 µg knihovna DNA byla štěpena s 0,5 nM nebo 50 nM Cas9-sgRNA složité (a), prodloužena do polohy 10 přesunutím protospacer cíl o 3 bp (b). Frekvence nukleotidů v každé poloze PAM byla nezávisle vypočtena pomocí matice polohové frekvence (PFM) a vykreslena jako WebLogo ., C míra štěpení superolovaných substrátů plazmidové DNA obsahujících mutace (znázorněné červeně)v sekvenci GTCCGAA PAM. Všechny datové body jsou střední hodnoty z ≥3 nezávislých experimentů. Chybové úsečky jsou uvedeny jako S. d. Run-off sekvenování z obou sense a anti-sense směrech plazmidové DNA štěpí Blat Cas9
Od Blat Cas9 může přijmout jakékoli základny v prvních třech pozic jeho PAM sekvence (Obr. 4A), distanční T1 byl posunut o tři nukleotidy ve směru 5′, aby bylo možné identifikaci PAM rozšířit ze 7 na 10 bp., Posunutý T1 distanční, T1-3 (AAACGCUAAAGAGGAAGAGG), bylo začleněno do Blat „přímé“ sgRNA a PAM identifikace byla provedena, jak je popsáno dříve pro Spy, Sth3, Sth1, a Blat Cas9 proteiny. Analýza preferencí PAM odhalila specifičnost PAM pro Blat Cas9 může být rozšířena na pozici 8, kde existuje mírná preference pro další A(obr. 4b).
PAM specificita pro Blat Cas9 byl potvrzen generováním plasmidy obsahují mutace v nejvíce zachované zbytky PAM (Obr. 4c)., Nahrazení nukleotidu C v pozici 5 zrušilo štěpení plazmidové DNA potvrzující jeho klíčovou roli v rozpoznávání PAM Blat Cas9. Nahrazení nukleotidů na pozicích 7 a 8 se významně snížil (43× a 12×, respektive) štěpení rychlost supercoiled plasmidu také naznačuje význam těchto nukleotidů v Blat Cas9 PAM uznání.,
identifikovat cílové DNA štěpení pozice pro Blat Cas9 bílkovin, plasmid obsahující 20 bp regionu odpovídající distanční T1 následovaný PAM sekvence, GTCCCGAA, spadající do PAM konsensus pro Blat Cas9, NNNNCNDD, byl vytvořen a stravitelné s Blat Cas9-guide RNA ribonucleoprotein komplexu. Přímé sekvenování DNA bylo použito k určení konců lineární molekuly DNA generované komplexem Blat Cas9 RNP. Výsledky sekvence potvrdily, že k štěpení PLAZMIDOVÉ DNA došlo v protospaceru 3 nt 5′ sekvence PAM (obr., 4d) podobně jako u proteinů Spy, Sth3 a Sth1 Cas9 .
V planta editování genomu pomocí Blat Cas9 a sgRNA
Následující objasnění sgRNA a PAM preference pro Blat Cas9, kukuřice optimalizované Cas9 a sgRNA výraz kazety byly vytvořeny v planta testování jak bylo již dříve popsáno u S. pyogenes cas9 gene a sgRNA . Krátce, Blat cas9 gene kukuřice kodonu optimalizované a intron 2 brambory ST-LSI gen byl vložen k narušení exprese v E. coli a usnadnit optimální sestřih v planta (Další soubor 1: Obrázek S12)., Jaderná lokalizace Blat Cas9 bílkovin v kukuřici buněk bylo usnadněno přídavkem obou amino a karboxylové terminální jaderné místech signály, SV40 (MAPKKKRKV) a Agrobacterium tumefaciens VirD2 (KRPRDRHDGELGGRKRAR), respektive (Další soubor 1: Obrázek S12). Na Blat cas9 gene byl konstitutivně exprimovaných v rostlinných buňkách propojením optimalizované cas9, aby kukuřice Ubiquitin promotér a pinII terminátor v plasmid DNA vektoru., Poskytují efektivní sgRNA výraz v kukuřici buněk, kukuřice U6 polymeráza III promotor a terminátor (TTTTTTTT), byly izolované a taveného na 5′ a 3′ koncích upravené Blat sgRNA kódování sekvence DNA, respektive (Další soubor 1: Obrázek S13). Upravený Blat sgRNA obsahoval dvě změny od používaných v in vitro; T G změna na pozici 99 a T C změna na pozici 157 sgRNA (Další soubor 1: Obrázek S13). Změny byly zavedeny k odstranění potenciálních předčasných terminačních signálů U6 polymerázy III v Blat sgRNA., Změny, kde představil mají minimální vliv na sekundární strukturu sgRNA oproti verzi používané v in vitro studiích (údaje nejsou uvedeny).
přesně porovnat mutační účinnosti vyplývající z nedokonalé non-homologní end-joining (NHEJ) opravy DNA double-strand breaks (DSBs) vyplývající z Spy a Blat Cas9 štěpení, protospacer stejné genomické cílové lokality byly vybrány na základě identifikace cílů, s Spy a Blat Cas9 kompatibilní PAMs, NGGNCNDD., Identické distanční sekvence byly vybrány pro Blat a Spy Cas9 zachycením sekvence 18 až 21 nt bezprostředně před PAM. K zajištění optimální U6 polymeráza III projevu a ne zavádět nesoulad v sgRNA distanční, všechny cílové sekvence byly vybrány tak, aby přirozeně ukončit v G na svém 5′ konci. Cíle byly identifikovány a vybrány v exonu 1 a 4 genu plodnosti kukuřice Ms45 a v oblasti před genem kukuřice liguleless-1.,
mutační aktivita Blat Cas9 u kukuřice byla zkoumána biolisticky transformujícími 10denní nezralá embrya kukuřice (IMEs) s DNA vektory obsahujícími geny cas9 a sgRNA. Blat a ekvivalentní Spy Cas9 a sgrna expresní vektory byly nezávisle zavedeny do kukuřice Hi – Type II IMEs transformací částic podobné tomu, který je popsán v. Od částicovou zbraň transformace může být velmi variabilní, visual marker DNA výraz kazety, Ds-Červené, byl také co-dodáván s Cas9 a sgRNA vektory exprese na pomoc při výběru rovnoměrně transformována IMEs., Celkem byly provedeny tři replikáty transformace na 60-90 IMEs a 20-30 z nejvíce rovnoměrně transformovaných IMEs z každé replikace bylo sklizeno 3 dny po transformaci. Celková genomická DNA byla extrahována a oblast obklopující cílové místo byla zesílena PCR a amplikony sekvenovanými do hloubky čtení přesahující 300 000. Výsledný čte byly vyšetřeny na přítomnost mutací v očekávaném místě štěpení ve srovnání s kontrolní experimenty, kde sgRNA DNA výraz kazeta byla vynechána z transformace. Jak je znázorněno na obr., 5A, mutace byly pozorovány v očekávaném místě štěpení pro Blat Cas9 s nejrozšířenějšími typy mutací, které jsou vkládáním nebo delecí jednoho základního páru. Podobné opravné vzory byly také pozorovány u proteinu Spy Cas9 (další soubor 1: obrázek S14 a ). Mutační aktivita pro Blat Cas9 byla robustní na dvou ze tří testovaných míst a překročila aktivitu Spy Cas9 na cílovém místě Ms45 exon 4 přibližně o 30% (obr. 5b).
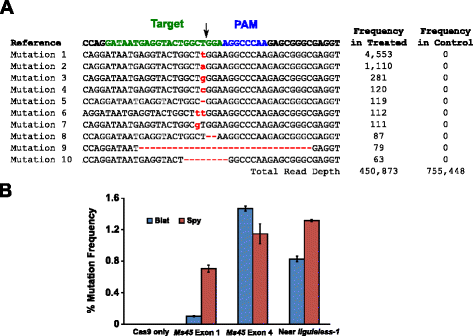
Brevibacillus laterosporus Cas9 podporuje NHEJ mutace v kukuřici. Top 10 nejrozšířenějších typů mutací NHEJ detekovaných pomocí Blat Cas9 v exonu 4 genu Ms45. Černá šipka označuje očekávané místo štěpení; mutace jsou zvýrazněny červeně; malá písmena označují vložení; “ – “ označuje vymazání. B porovnání četností mutací Spy a Blat Cas9 NHEJ na třech protospacerových identických cílových místech v kukuřici. Nhej mutace byly detekovány hlubokým sekvenováním 3 dny po transformaci., Chybové tyče představují sem, N = 3 přeměny částic. Cas9 je pouze negativní kontroly a představuje průměrnou (ve všech třech cílových míst) pozadí frekvence mutací vyplývající z PCR amplifikace a sekvenování