Suunnittelu ja rakentaminen satunnaistettu kirjastot leimaamisen Cas9 PAM-asetukset
PAM kirjastot sisältävät satunnaistettu DNA-sekvenssit välittömästi alavirtaan DNA-sekvenssi, joka täydentää spacer opas-RNA oli tuottaa ja käyttää empiirisesti määrittää PAM tunnustaminen II-Tyypin Cas9 endonucleases (Fig. 1)., Opas-RNA spacer kohde järjestys on kiinteä, satunnaistettu emäkset toimivat substraatti suoraan lukea ulos Cas9 endonuclease PAM spesifisyys. Satunnaistettu sekvenssit olivat tuodaan plasmidi-DNA-vektorin PAM alue protospacer kohdesekvenssin osoittaa täydellinen homology opas-RNA spacer T1 (CGCUAAAGAGGAAGAGGACA). Kaksi kirjastot koko kasvaa ja monimutkaisuus viisi satunnaistettu emäsparin (1,024 mahdolliset PAM yhdistelmät) seitsemän satunnaistettu emäsparin (16,384 mahdolliset PAM yhdistelmiä) on luotu., Satunnaistaminen 5 bp-kirjasto otettiin käyttöön synteesin kautta yhden oligonukleotidi, jotka sisältävät viisi satunnaisesti jäämiä. Single-pulaan oligonukleotidi oli muunnetaan double-stranded malli PCR: llä (Lisää tiedosto 1: Kuva S1A), kloonattu osaksi plasmidia vektori (Lisää tiedosto 1: Kuva S1B) ja transformoitiin E. coli kuvattu Menetelmät-osiossa., Optimaalisen satunnaisuus 7 bp PAM library, koko ja monimutkaisuus kirjaston väheni syntetisointi neljä oligonucleotides, joista jokainen sisältää kuusi satunnaisia jäämiä sekä seitsemäs kiinteä jäännös, joka käsittää G, C, A tai T, vastaavasti. Kunkin neljän oligonukleotidit olivat erikseen muunnetaan double-stranded DNA, kloonattu osaksi vektori pTZ57R/T on kuvattu Menetelmät-osiossa ja transformoitiin E. coli kuvattu 5 bp kirjasto., Kun muutos, plasmidi-DNA: ta otettiin talteen ja yhdistetään kustakin neljä 6 bp PAM kirjastot tuottaa satunnaistettu 7 bp PAM library sisältää 16,384 mahdollista PAM yhdistelmiä. Sekä kirjastojen sisällyttäminen satunnaisuus oli vahvistanut syvä sekvensointi; tutkimalla nukleotidin koostumus kummassakin asennossa PAM alueella käyttäen aseman taajuus matriisi (PFM) (Menetelmät-osiossa ja ) (Lisää tiedosto 1: Kuva S2A ja B)., Jakelu ja taajuus kunkin PAM järjestyksessä 5 bp ja 7 bp satunnaistettu PAM library on esitetty Tiedostojen 1: Luvut S3 ja S4, vastaavasti.
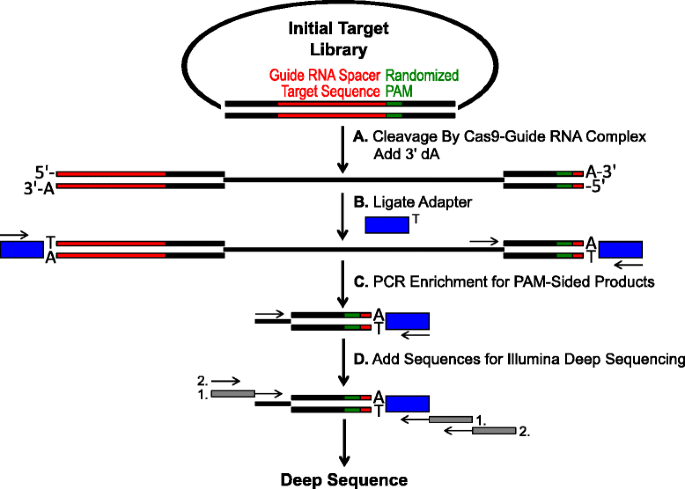
Kaavio tunnistamiseksi PAM mieltymysten mukaan Cas9 pilkkominen in vitro. aluksi plasmidi kirjasto, jossa on satunnaistettu PAM (vihreä laatikko) on hajonnut kanssa Cas9 monimutkainen ja 3′ dA ulokkeita lisätään. B-Adapterit, joissa on 3′ dT overhang (blue box), sidotaan katkaisutuotteen molempiin päihin., C Primers hyödynnetään rikastuttaa Pam-puolinen cleaved tuotteita PCR., d sen Jälkeen PCR-rikastamiseen, DNA-fragmentit puhdistettiin ja Illumina-yhteensopiva ankkurit ja viivakoodit ovat ’pyrstö-on’ läpi kaksi kierrosta PCR (harmaat laatikot) ja Illumina syvä sekvensoitiin
Leimaamisen Cas9 PAM-asetukset
satunnaistettu PAM kirjastot kuvattu edellisessä jaksossa tehtiin in vitro ruoansulatusta eri pitoisuuksia yhdistelmä-Cas9 proteiinia esiladattu opas-RNA jotta määritys Cas9 endonuclease PAM mieltymykset annoksesta riippuvalla tavalla., Jälkeen ruoansulatusta kanssa Cas9-opas-RNA ribonucleoprotein (RNP) komplekseja, PAM järjestyksessä yhdistelmiä satunnaistettu PAM-kirjasto, joka tukee pilkkominen olivat vangiksi ligaatio-adapterit vapaa-päät plasmidi-DNA-molekyylit pilkotaan, jonka Cas9-opas-RNA-kompleksi (Kuva. 1 A ja b). Edistää tehokkaita sitomiseen ja kaapata halkaistut päät, tylppä-päättyi kaksinkertaisen-pulaan DNA leikkaa syntyy Cas9 endonucleases oli muutettu sisältää 3′ dA ylitys ja adapterit muutettiin sisältää täydentävä 3′ dT ylitys., Tuottaa riittäviä määriä DNA: n sekvensointi, DNA-fragmentit kätkeminen PAM järjestyksessä tukeminen pilkkominen olivat PCR-monistetaan käyttämällä pohjamaali sovitin ja toinen suoraan vieressä PAM alueella (Kuva. 1c). Tuloksena PCR täydennetty Cas9 Pam kirjastot muutettiin ampli-seq malleja (kuva. 1D) ja yksilukuinen syvä sekvensoitu adapteri-puolella amplicon., Varmistaa riittävä kattavuus, Cas9 PAM kirjastot sekvensoitiin syvyys vähintään viisi kertaa suurempi kuin monimuotoisuuden ensimmäinen satunnaistettu PAM library (5,120 ja 81,920 lukee 5 ja 7 bp PAM satunnaistettu kirjastot, vastaavasti). PAM sekvenssit määritettiin saadusta järjestyksessä tiedot valitsemalla vain ne lukee, jotka sisältävät täydellinen 12 nt järjestyksessä ottelu reunustavat kummallakin puolella 5 tai 7 nt PAM järjestyksessä (riippuen satunnaistettu PAM library käytössä); syömällä vain ne, PAM sekvenssit, jotka johtuvat täydellinen Cas9-opas-RNA kohde-sivuston tunnustaminen ja pilkkominen., Kompensoimaan luonnostaan harhaa ensimmäinen satunnaistettu PAM kirjastot, taajuus kunkin PAM sekvenssi oli normalisoitu taajuus alkaa library. Koska tässä kuvattu määritys kaappaa suoraan Cas9 cleavable PAM sekvenssit, probabilistinen mallinnus käytettiin laskemaan PAM konsensus kunkin Cas9 proteiinia. Tämä toteutettiin arvioimalla todennäköisyys löytää kunkin nukleotidin (G, C, A tai T) kummassakin asennossa PAM järjestyksessä itsenäisesti käyttäen aseman taajuus matriisi (PFM) (Menetelmät-osiossa ja )., Tuloksena olevat todennäköisyydet visualisoitiin tämän jälkeen Weblogoksi .
tutkittaessa väärien positiivien taipumusta määrityksessä Cas9 RNP-kompleksien lisääminen ruuansulatusvaiheeseen jätettiin pois (Kuva. 1A) ja määritys suoritettiin PCR: n rikastusaskeleella (Kuva. 1c). Kuten näkyy Tiedostojen 1: Kuva S5A, ei vahvistus tuotteiden havaittu absense Cas9-opas-RNA-komplekseja. Näin ollen, osoittaa, että esiintyvyys vääriä positiivisia on alhainen eikä merkittävästi edistää tuloksia määrityksen.,
PAM mieltymykset Streptococcus pyogenes ja Streptococcus thermophilus (CRISPR3 ja CRISPR1 systems) – Cas9 proteiineja
jotta voidaan vahvistaa assay, PAM mieltymykset Streptococcus pyogenes (Spy) ja Streptococcus thermophilus CRISPR3 (Sth3) Cas9 proteiineja, joiden PAM järjestyksessä vaatimus on aiemmin todettu , tutkittiin. In vitro-tiivistelmät tehtiin 1 µg (5.6 nM) 5 bp satunnaistettu PAM library kaksi pitoisuuksia, 0,5 ja 50 nM -, pre-koottu Vakooja tai Sth3 Cas9 proteiinia, crRNA, ja tracrRNA RNP komplekseja 1 s show 100 µL reaktion tilavuus., Perustuu niiden taajuus 5 bp satunnaistettu PAM library, Vakooja ja Sth3 Cas9 PAM sekvenssit (NGG-ja NGGNG, vastaavasti) olivat lopulliset pitoisuudet 0,40 nM ja 0,11 nM ruoansulatusta, vastaavasti. Satunnaistetun Pam-kirjaston jäsenet, jotka sisälsivät Pam-sekvenssejä, jotka tukivat pilkkoutumista, vangittiin ja tunnistettiin edellisessä jaksossa kuvatulla tavalla. Kuin negatiivinen kontrolli alkaa katkaisemattomat satunnaistettu PAM library oli kohteena sekvensointi ja PFM-analyysin rinnalla kirjastot alttiina Cas9 RNP komplekseja., Kuten näkyy Tiedostojen 1: Kuva S5B ja C, ei järjestyksessä mieltymysten olemassa ilman Cas9 RNP-kompleksi ruoansulatusta kuten ilmeistä lähes täydellinen jakelu kunkin nukleotidin kummassakin asennossa PAMIN PFM-taulukko ja puute informatiivinen sisältö WebLogo valvontaa. Tämä on stark constrastissa Figin kanssa. 2a ja b, joka havainnollistaa koostumus sekvenssien johdettu kirjastot pilkottu Spy ja Sth3 Cas9 RNP komplekseja. PFM-johdettujen Weblogojen tutkiminen (Kuva., 2 a ja b) myös paljastaa läsnäolo kanoninen PAM mieltymykset Vakooja ja Sth3 Cas9 proteiineja, NGG-ja NGGNG , vastaavasti. Vaikka PAM mieltymykset todettu, Vakooja ja Sth3 Cas9 proteiinien on havaittu sekä 0,5 nM ja 50 nM sulattaa, on yleinen laajentaa vuonna spesifisyys alle 50 nM sulattaa ehtoja. Tämä on selvimmin asennossa 2 Vakooja Cas9 proteiinia, jossa taajuus ei-kanoninen jäännös kasvaa dramaattisesti (Kuva. 2 a)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
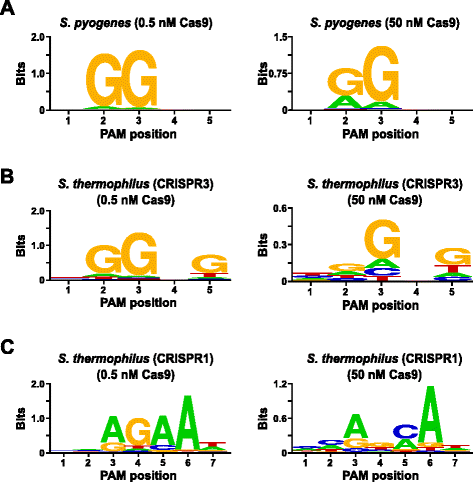
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., Taajuus nukleotidien kunkin PAMIN kanta oli itsenäisesti laskettu käyttäen aseman taajuus matriisi (PFM) ja piirretään WebLogo
Edelleen määritysmenetelmän validoinnin yhteydessä tehtiin tutkimalla PAM mieltymykset Streptococcus thermophilus CRISPR1 (Sth1) Cas9-proteiinia, jonka PAM spesifisyys on raportoitu pidentää ulos 7 bp . Käyttämällä 1 µg (5.6 nM) 7 bp satunnaistettu PAM library mallina, Sth1 Cas9-opas-RNA digestions tehtiin kaksi pitoisuutta, 0.5 nM ja 50 nM, ja RNP-kompleksi, kuten edellä on kuvattu., Verrokkeina, Vakooja ja Sth3 Cas9 RNP komplekseja käytettiin myös sulattaa 7 bp satunnaistettu PAM library mutta vain yhden, 0.5 nM, RNP-kompleksi keskittymistä. Perustuu taajuus 7 bp satunnaistettu PAM kirjasto, PAM sekvenssit aiemmin raportoitu Sth1 (NNAGAAW), Vakooja (NGG), ja Sth3 (NGGNG) olivat lopulliset pitoisuudet 0,01 nM, 0.22 nM, ja 0,05 nM, vastaavasti., Kuten näkyy Tiedostojen 1: Kuva S6A-ja B-PAM mieltymykset Vakooja ja Sth3 Cas9 proteiineja tuotetaan käyttämällä 7 bp library olivat lähes identtisiä valmistettu 5 bp kirjasto tarjoaa vahvaa näyttöä siitä, että toistettavuus määritys. PAM mieltymykset Sth1 Cas9 proteiinia myös lähellä toisiaan, että aiemmin on raportoitu, NNAGAAW , tällä 0.5 nM Cas9-opas-RNA-kompleksi pitoisuus (Fig. 2 C)., Samanlainen Vakooja ja Sth3 Cas9 proteiineja, Sth1 Cas9 pystyy pilkkomaan monipuolisempi joukko PAM sekvenssit reaktioita, jotka sisältävät korkeamman pitoisuuden Cas9-opas-RNA-kompleksi (50 nM), silmiinpistävin oli merkitty menetys G jäännös vaatimus asentoon 4, ja lähes yhtä suuri mieltymys C ja bp 5. asennossa (Kuva. 2 C). Tämä johti erilaiseen PAM-konsensukseen kuin pienempinä pitoisuuksina saatu konsensus.,
tutkia, onko PAM spesifisyys on riippumaton tyyppi opas-RNA: ta, yksi crRNA:tracrRNA tai sgRNA , Vakooja, Sth3, ja Sth1 Cas9 PAM mieltymykset tutkittiin käyttämällä binary Cas9 ja sgRNA RNP-kompleksi. Ruoansulatus suoritettiin yhdellä RNP-kompleksipitoisuudella, joka oli 0,5 nM, ja PAM-suositusanalyysi tehtiin edellä kuvatulla tavalla. Kuten näkyy Tiedostojen 1: Kuva S7A, B, ja C, PAM mieltymykset olivat lähes identtiset riippumatta siitä, millainen opas-RNA käytetty; joko crRNA:tracrRNA duplex tai sgRNA., Lisäksi vahvistaa, että PAM spesifisyys ei ole suuresti vaikuttanut koostumus kohde-DNA: n tai spacer sekvenssi, sekvenssi vastakkaisella puolella 5 tai 7 bp satunnaistettu kirjasto oli suunnattu pilkkominen eri spacer; T2-5 (UCUAGAUAGAUUACGAAUUC) 5 bp kirjasto-tai T2-7 (CCGGCGACGUUGGGUCAACU) 7 bp kirjasto. Vakooja ja Sth3 Cas9 proteiineja esiladattu sgRNAs kohdistaminen T2-sekvenssi oli tapana kuulustella 5 bp satunnaistettu PAM library vaikka Sth1 Cas9-T2 sgRNA komplekseja käytettiin sulattaa 7 bp satunnaistettu PAM library., PAMin mieltymykset analysoitiin edellä kuvatulla tavalla. PAM mieltymykset kaikille 3 Cas9 proteiinit olivat lähes identtiset riippumatta siitä, välike ja kohde-DNA: n sekvenssi (Lisää tiedosto 1: Kuva S8A -, B -, ja C).
Tunnistaminen sgRNA ja PAM mieltymykset Brevibacillus laterosporus Cas9 proteiinia
empiirisesti tutkia PAM mieltymykset Cas9-proteiinia, jonka PAM oli määrittelemätön, on uncharacterized Tyyppi II-C CRISPR-Cas locus päässä Brevibacillus laterosporus kanta SSP360D4 (Blat) tunnistettiin etsimällä sisäisen DuPont Pioneer tietokantojen Cas9 orthologues., Locus (noin 4.5 kb) sisälsi cas9-geeni, joka pystyy koodaus 1,092 polypeptidi, a CRISPR joukko, johon kuuluu seitsemän toista-spacer yksiköt vain alavirtaan cas9-geenin ja tracrRNA koodaus alueella sijaitsee ylävirtaan cas9-geenin osittainen homology, että CRISPR array toistaa (Kuva. 3 A). Toista ja välikappale pituus (36 ja 30 bp, vastaavasti) on samanlainen kuin muut Tyypin II CRISPR-Cas-järjestelmien kanssa viisi kahdeksan toistoja, joissa on 1 tai 2 bp-mutaatioita (Kuva. 3b ja lisätiedosto 1: kuva S9)., Muut tyypin II CRISPR-CAS-lokuksessa tyypillisesti esiintyvät geenit olivat joko typistettyjä (cas1) tai puuttuvia (Fig. 3 A).
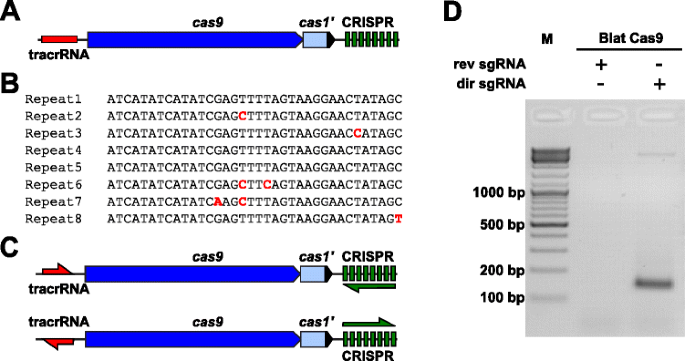
Tunnistaminen Tyypin II CRISPR-Cas-elementtejä Brevibacillus laterosporus SSP360D4 CRISPR-Cas-järjestelmä. yksi esimerkki genomien DNA-alueen Tyyppi II CRISPR-Cas-järjestelmän Brevibacillus laterosporus SSP360D4. b Vertailu Tyypin II CRISPR array toista sekvenssit tunnistettu Brevibacillus laterosporus SSP360D4., c ’suora’ ja ’käänteinen’ tracrRNA ja CRISPR array transkription skenaarioita Tyyppi II CRISPR-Cas-järjestelmän Brevibacillus laterosporus SSP360D4. d agaroosia geeli reaktion tuotteita, mikä osoittaa, että vain ”suora” sgRNA (dir sgRNA), mutta ei ’käänteinen’ sgRNA (rev sgRNA) tukea plasmidi kirjasto pilkkominen yhdessä Cas9 endonuclease peräisin Brevibacillus laterosporus SSP360D4
– opas-RNA vaatimus Blat Cas9 proteiini määritettiin luomalla kaksi sgRNA variantteja., Nämä vaihtoehdot oli luotu tili sekä mahdolliset järkeä tai anti-sense ilmaisun skenaarioita tracrRNA ja CRISPR array (Fig. 3c) ja käytetään tutkimaan, mikä lauseke skenaario tuki pilkkoutumista toimintaa Blat Cas9 satunnaistetussa PAM kirjasto. Yhden oppaan RNAs oli suunniteltu tunnistamalla ensin rajat oletetun tracrRNA molekyylejä analysoimalla alueilla, jotka olivat osittain toisiaan täydentäviä 22 nt 5′ päähän toista (anti-repeat)., Seuraavaksi, määrittää 3′ loppuun tracrRNA, mahdolliset toissijaiset rakenteet ja terminaattorit käytettiin ennustaa alueen irtisanominen loppupään fragmentti. Tämä oli suoritettu seulonta läsnäolo Rho riippumattomia-kuten irtisanominen sekvenssejä DNA: ta ympäröivä anti-toista samanlainen kuin on kuvattu Karvelis et al. , muuntamalla ympäröivän DNA: n RNA-sekvenssiksi ja tutkimalla tuloksena olevia rakenteita UNAfold-menetelmällä ., Tuloksena sgRNAs oli tarkoitus sisältää T7-polymeraasi transkriptio aloittamista tunnustamista signaalin 5′ pää seurasi 20 nt kohteen tunnistus järjestyksessä, 16 nt of crRNA toista, 4 nt self-taitto hiusneula silmukka, ja anti-toista järjestyksessä täydentää toista alueen crRNA jonka jälkeen loput 3 osa oletetun tracrRNA. Sgrna-muunnos, joka sisältää putatiivisen tracrRNA: n transkriboituna samaan suuntaan kuin cas9-geeni (Fig. 3c) on ilmaisua ”suora” sgRNA, kun sgRNA sisältää tracrRNA puhtaaksi vastakkaiseen suuntaan ’käänteinen’ sgRNA., Viisikymmentä nM Blat Cas9 sgRNA RNP-kompleksi, valmiiksi ladattu joko ”suora” tai ’taaksepäin’ sgRNAs, vastaavasti, olivat inkuboitiin 1 µg (5.6 nM) 7 bp satunnaistettu PAM library. Kirjaston digestion ja 3′ dA overhangsin lisäämisen jälkeen adapterit sidottiin ja pilkkoutumistuotteet vahvistettiin PCR: llä (Kuva. 1). Analyysi reaktion tuotteet agaroosia geeli elektroforeesi paljasti, että ”suora” sgRNA, mutta ei ’käänteinen’ sgRNA tuettu plasmidi kirjasto pilkkominen (Fig. 3D). ”Suoran” sgRNA: n sekvenssi ja ennustettu toissijainen rakenne esitetään lisätiedostossa 1: kuva S10.,
Kun määritetään sopiva opas-RNA: ta varten Blat Cas9, PAM tunnistaminen suoritettiin samalla tavalla kuin edellä on kuvattu Vakooja, Sth3, ja Sth1 Cas9 proteiineja vastaan 7 bp satunnaistettu PAM-kirjaston kanssa kaksi pitoisuutta, 0,5 ja 50 nM -, pre-koottu Blat Cas9 ’suora’ sgRNA RNP-kompleksi. Kuten kuvassa. 4a, PFM WebLogo PAM yhteisymmärrykseen Blat Cas9 proteiini alle 0,5 nM sulattaa ehtoja oli NNNNCND (N = G, C, A tai T, D = A, G tai T), jolla on vahva mieltymys C asennossa 5 PAM järjestyksessä., Maltillinen ratkaisu oli havaittu kannan 7 ja hieman mieltymykset C tai T-asennossa 4 ja G, C, tai yli T-asennossa 6 havaittiin myös, kun tarkasti tutkii PFM-taulukko (Lisää tiedosto 1: Kuva S11). Samoin kuin Spy -, Sth3-ja Sth1 Cas9-proteiineissa PAMin spesifisyys laajenee Cas9-sgRNA-kompleksin pitoisuuden kasvaessa. Tämä on selvimmin 5. asennossa, jossa suurempi osa PAM sekvenssit, jotka sisältävät jäämiä tukea pilkkominen 50 nM verrattuna 0,5 nM sulattaa ehtoja.
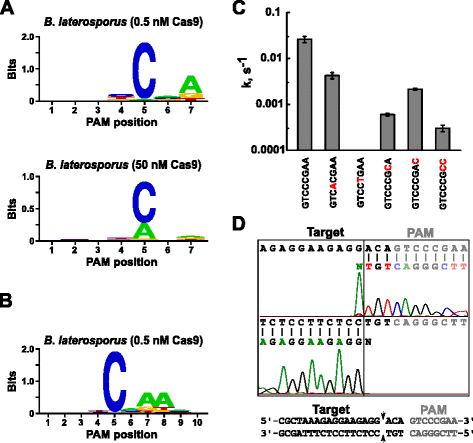
PAM mieltymykset ja pilkkominen kannat Brevibacillus laterosporus SSP360D4 (Blat) Cas9-entsyymiä. Blat Cas9 PAM mieltymykset, kun 1 µg DNA-kirjasto oli halkaistut 0,5 nM tai 50 nM Cas9-sgRNA monimutkainen (a), laajennettu asentoon 10 siirtämällä protospacer tavoite 3 bp (b). Nukleotidien esiintymistiheys kussakin PAM-asennossa laskettiin itsenäisesti käyttäen positiotaajuusmatriisia (PFM) ja piirrettiin Weblogoksi ., C ylijääneiden plasmidi-DNA-substraattien Pilkkoutumisnopeudet, jotka sisältävät mutaatioita (punaisella) GTCCGAA Pam-sekvenssissä. Kaikki datapisteet ovat keskiarvoja ≥3 riippumattomasta kokeesta. Virhepalkit ovat antaneet S. D. d Run-off-sekvensointi sekä tunne-ja anti-sense suuntaan plasmidi-DNA pilkotaan kanssa Blat Cas9
Koska Blat Cas9 voi hyväksyä mitään pohja kolmen ensimmäisen kantoja sen PAM järjestyksessä (Kuva. 4a), välike T1 oli siirtynyt kolmen nukleotidin 5′ suuntaan, jotta PAM tunnistaminen laajennetaan 7 10 bp., Siirtynyt T1 spacer, T1-3 (AAACGCUAAAGAGGAAGAGG), otettiin osaksi Blat ’suora’ sgRNA ja PAM tunnistaminen suoritettiin kuten aiemmin on kuvattu Vakooja, Sth3, Sth1, ja Blat Cas9 proteiineja. PAM etusija analyysi paljasti PAM spesifisyys Blat Cas9 voidaan pidentää ulos asentoon 8, jossa on kohtalaista parempana ylimääräinen A (Fig. 4 B).
PAM spesifisyys Blat Cas9 vahvistettiin tuottamalla plasmidit sisältävät mutaatioita kaikkein säilytetty jäämät PAM (Fig. 4 C)., C-nukleotidin korvaaminen kohdassa 5 poisti plasmidin DNA-pilkkoutumisen vahvistaen sen keskeisen roolin Blat Cas9 PAM-tunnistuksessa. Korvaaminen nukleotidien kohdissa 7 ja 8 huomattavasti (43× 12×, vastaavasti) pilkkominen määrä supercoiled plasmidi myös osoittaa, miten tärkeää näiden nukleotidien Blat Cas9 PAM tunnustamista.,
tunnistaa DNA tavoitteen pilkkominen kannat Blat Cas9 proteiinia, plasmidi, joka sisältää 20 bp alueen vastaavat spacer T1 seuraa PAM järjestyksessä, GTCCCGAA, jotka kuuluvat PAM yksimielisyys Blat Cas9, NNNNCNDD, oli luotu ja pilkottu Blat Cas9-opas-RNA ribonucleoprotein monimutkainen. Suoran DNA-sekvensoinnin avulla määritettiin blat Cas9 RNP-kompleksin tuottaman lineaarisen DNA-molekyylin päät. Sekvenssitulokset vahvistivat, että plasmidin DNA-pilkkoutuminen tapahtui Pam-sekvenssin protospacer 3 nt 5′: ssä (kuva., 4d )samanlainen kuin Spy -, Sth3-ja Sth1 Cas9-proteiineissa.
planta genomin editointi käyttäen Blat Cas9 ja sgRNA
Seuraavat selvittäminen sgRNA ja PAM mieltymykset Blat Cas9 -, maissi-optimoitu Cas9 ja sgRNA ilmaisun kasetteja kertyi in planta testaus kuten aiemmin on kuvattu S. pyogenes-cas9-geenin ja sgRNA . Lyhyesti, Blat cas9-geeni oli maissin kodonissa optimoitu ja introni 2 peruna-ST-LSI-geeni oli asetettu häiritä ilmaisun E. coli ja helpottaa optimaalinen liitos planta (Lisää tiedosto 1: Kuva S12)., Ydinvoiman lokalisointi Blat Cas9 proteiini maissi solujen helpotti lisäksi sekä amino-ja karboksyyli-terminaali ydinvoiman paikoissa signaaleja, SV40 (MAPKKKRKV) ja Agrobacterium tumafaciensilla VirD2 (KRPRDRHDGELGGRKRAR), vastaavasti (Lisää tiedosto 1: Kuva S12). Blat cas9-geeni ilmaistiin kasvisoluissa konstitutiivisesti yhdistämällä optimoitu cas9 maissin Ubikitiinipromoottoriin ja pinII-terminaattori plasmidin DNA-vektoriin., Antaa tehokkaan sgRNA ilmaisun maissin soluja, maissi U6-polymeraasi III promoottori ja terminator (TTTTTTTT) eristettiin ja sulatettu 5′ – ja 3′ – päät muutettu Blat sgRNA koodaus DNA-sekvenssi, vastaavasti (Lisää tiedosto 1: Kuva S13). Muutettu Blat sgRNA sisälsi kaksi muutosta siitä, että käytetään in vitro-tutkimukset; T G muutos asennossa 99 ja T-C muutos asennossa 157 sgRNA (Lisää tiedosto 1: Kuva S13). Muutokset otettiin käyttöön mahdollisten ennenaikaisten U6-polymeraasi III-päättymissignaalien poistamiseksi Blat sgRNA: sta., Muutokset, joilla on vähäinen vaikutus sgRNA: n toissijaiseen rakenteeseen verrattuna in vitro-tutkimuksissa käytettyyn versioon (tietoja ei ole esitetty).
tarkasti verrata mutaation tehokkuutta, jotka johtuvat epätäydellinen ei-homologisia end-liittymällä (NHEJ) korjaus DNA-double-säikeen katkoksia (DSBs), jotka johtuvat Vakooja ja Blat Cas9 pilkkominen, protospacer identtinen genomi kohde sivustot valittiin tunnistamalla tavoitteita, Vakooja ja Blat Cas9-yhteensopiva Pamit, NGGNCNDD., Identtiset spacer sekvenssit valittiin Blat ja Spy Cas9 syömällä 18-21 nt sekvenssi välittömästi ennen PAM. Optimaalisen U6-polymeraasi III ilmaisun ja käyttöön epäsuhta sisällä sgRNA spacer, kaikki kohde-sekvenssit valittiin luonnollisesti lopettaa G niiden 5′ pää. Tavoitteet oli tunnistettu ja valittu eksonin 1 ja 4 maissin hedelmällisyyttä geeni Ms45 ja alueella ylävirtaan maissin liguleless-1-geenin.,
– mutaation toimintaa Blat Cas9 maissin tarkasteli biolistically muuttamassa 10 päivän ikäinen epäkypsä maissin alkioita (Ime), jossa DNA-vektoreita, jotka sisältävät cas9 ja sgRNA geenit. Blat ja vastaa Spy Cas9 ja sgRNA ilmaisun vektorit olivat itsenäisesti tuodaan maissia Hi-Tyyppi II Ime, jonka hiukkanen ase muutosta kuvatun kaltaista . Koska hiukkanen ase muutos voi olla erittäin vaihteleva, visuaalinen merkki DNA-ilmaisun kasetti, Ds-Punainen, oli myös co-toimitetaan Cas9 ja sgRNA ilmaisun vektorit tuen valinnassa tasaisesti muuttunut Ime., Yhteensä kolme muutosta rinnakkaisnäytteitä tehtiin 60-90 Ime ja 20-30 kaikkein tasaisesti muuttunut Ime kustakin rinnakkaisnäytteestä oli korjattu 3 päivän kuluttua muutosta. Koko genomista DNA uutettiin, ja ympäröivällä alueella kohde-sivuston monistettiin PCR: llä ja amplicons sekvensoitiin voit lukea syvyys on yli 300 000. Tuloksena lukee tutkittiin läsnäolo mutaatioita odotettavissa sivuston pilkkominen vertailun valvoa kokeita, joissa sgRNA DNA ilmaisun kasetti oli pois muutosta. Kuten kuvassa., 5a, mutaatioita havaittiin odotettavissa sivuston rintoja Blat Cas9 kanssa yleisin tyyppisiä mutaatioita on yhden emäsparin lisäyksiä tai poistoja. Samanlaisia korjauskuvioita havaittiin myös Spy Cas9-proteiinille (lisätiedosto 1: kuva S14 ja ). Sen mutaation toimintaa Blat Cas9 oli vankka kaksi kolmesta sivustoja testattu ja ylitti Spy Cas9 klo Ms45 eksonin 4 kohde-sivuston mukaan noin 30 % (Kuva. 5 b).
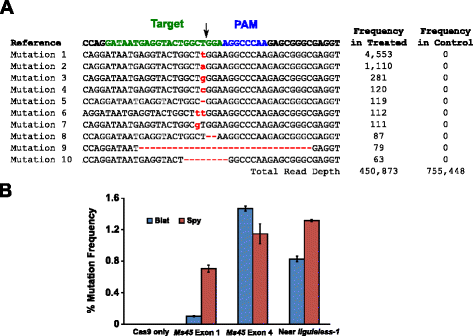
Brevibacillus laterosporus Cas9 edistää NHEJ mutaatioita maissia. Top 10 eniten vallalla erilaisia NHEJ mutaatioita havaittu Blat Cas9 eksonin 4 Ms45-geeni. Musta nuoli osoittaa odotetun pilkkoutumispaikan; mutaatiot korostuvat punaisina; alempi kirjasin osoittaa lisäyksen; ” – ” tarkoittaa poistoa. b Vertailu Vakooja ja Blat Cas9 NHEJ mutaatio taajuuksia kolme protospacer sama tavoite sivustoja maissia. NHEJ-mutaatiot havaittiin syväsekvensoimalla 3 päivää muodonmuutoksen jälkeen., Virhepalkit kuvaavat keskiarvon keskivirhe, n = 3 hiukkanen ase muutoksia. Cas9 vain on negatiivinen kontrolli ja edustaa keskimääräistä (kaikkien kolmen tavoitteen sivustoja) tausta taajuus mutaatiot johtuvat PCR-monistus ja sekvensointi