Vaikka haihtuvien yhdisteiden kanssa-agonistin toimintaa on seulottu vastaan ORs, tiettyyn kemiallinen vahva agonistin vaikutus hyttysiä, 2-(4-Etyyli-5-(pyridin-3-yyli)-4H-1,2,4-triatsol-3-ylthio)-N-(4-ethylphenyl)asetamidi (VUAA1), on avannut tutkimuksen (Jones et al., 2011; Taylor ym., 2012). Myöhemmät tutkimukset Taylor et al., (2012) esittivät todisteita siitä, VUAA1 johdettuja kemikaaleja, kuten VUAA4, voi lisätä sen agonistinen vaikutus on 10-kertainen ORco A. gambiae, H. virescens, ja Harpegnathos saltator. Mielenkiintoista on, että kirjoittajat kertoivat, että kaikki amide-substituenttien muutokset aiheuttavat täydellisen agonistitoiminnan menetyksen. Näin saadaan hyödyllistä tietoa agonistien rakenteellisista vaatimuksista ja vuaa-analogien ja ORs-analogien rakenteellisesta ja aktiivisesta suhteesta. Lopuksi, huolimatta enhanced-agonistin toimintaa VUAA kemikaaleja, sen suhteellisen korkea molekyylipaino (367.47 g mol−1 VUAA1) vs., haihtuvien agonisteilla, kuten bombykol (238.42 g mol−1), joka tekee suoran haihtuvien toimitus VUAA jotain ei ole mahdollista. Tässä mielessä pienempien rakenteellisten analogien etsiminen on mielenkiintoinen tutkimuksen painopiste.
Vihamielisyys Päälle ORco-Alayksikkö
Yhdessä tutkimuksessa VUAA-liittyvät analogit, jotka voivat toimia yhtä vahva agonistit, tukos ORco, jonka antagonisteja on myös syntynyt opas semiokemikaalien ja torjunta-suunnittelu., Näin ollen rakenteellinen analogi VUAA1, VU0183254 (2-(4-Etyyli-5-furan-2-yl-4H-triatsol-3-ylsulfanyl)-1-phenothiazin-10-yl-ethanone), oli todettu estävän ORco vastaus, joka toimii allosteerinen modulaattori A. gambiae ja häiritsevät tunnustaminen-agonistit, kuten eugenoli monimutkainen OR65/ORco (Jones et al., 2012). Myös muita vua-rakenteellisia analogeja on raportoitu antagonisteina. Esimerkkinä on N-,2-substituoitu triazolothioacetamide yhdisteitä OLC3 ja OLC12, joka häiritsee ORco vastausta samalla tavalla C. quinquefasciatus, A. gambiae, D. melanogaster ja O., nubilalis, mikä viittaa säilytetty sitova sivusto ORco (Chen ja Luetje, 2012). Ottaen huomioon, esto ORco lupaava strategia häiritä käyttäytymistä hyönteisiä, näyttää siltä, että myöhemmin olisi pyrittävä yhdisteiden kanssa pienempi molekyylipaino kuin VUAA-johdettu antagonistit. Esimerkiksi, OX1a (232 g mol−1), tryptamiini (160.22 g mol−1) ja isopropyyli cinnamate (190.24 g mol−1) ilmoitettiin ovat antagonisti vaikutus ORco (Chen ja Luetje, 2013, 2014; Tsitoura et al., 2015), jonka molekyylipaino on noin puolet tai vähemmän kuin VUAA1., Kuitenkin tulevaisuudessa käyttää näitä salpaajia, tulisi tutkittu huolellisesti, koska tukos säilytetty ORco voi vaikuttaa paitsi haitallisia hyönteisiä, mutta myös hyötyä niistä.
Lisäksi antagonisti vaikutus tutkittiin in vitro, todisteita käyttäytymisen tasolla tukee ajatusta siitä, että rakenteellisia analogeja feromonit voi toimia antagonistit. Esimerkiksi Sellanes et al., (2010) raportoi esto seksuaalisen vasteen hunajameloni koi Cryptoblabes gnidiella kun rakenneanalogit, (Z)-9-tetradecenyl formiaatit ja (Z)-11-hexadecenyl formaatti, lisättiin synteettistä sukupuoli feromoni, (Z)-11-hexadecenal ja (Z)-13-octadecenal, tuulitunnelikokeissa. Tämä feromoni antagonisti vaikutus oli myöhemmin vahvistaa kentän määrityksiin, joissa ansastusta C. gnidiella miehillä väheni annoksesta riippuvaa mallia. Feromoniantagonismia on raportoitu myös B. mandarinalle, joka on B. Morin (Daimon et al., 2012)., Niiden tulokset tukevat bombykol kuten sukupuoli feromoni, ja bombykal ja bombykyl asetaatti kuin antagonistit, joka on vahvasti estetty vetovoima urokset kentän sukupuoli feromoni bombykol. Viime aikoina on raportoitu näyttöä feromoniantagonismista kuonon Herpetogramma submarginalella. Kun (Z)-13-hexadecenol oli lisätty sen sukupuoli feromoni, (Z)-13-hexadecenyl asetaatti, merkittävästi vähentynyt määrä miehiä jää kenttään (Yan et al., 2015)., Feromoni vihamielisyys näyttää perustuu eroja kemiallinen funktionaalisen ryhmän, kuten alkoholeja, aldehydejä ja estereitä riippuen hyönteisten lajeja. Kuitenkin, antagonisti vaikutus näiden rakenneanalogit ei ehkä johtuu ORco esto, mutta spesifisyys ORx antagonistien. Tuore tutkimus viittaa siihen, että OR16 ja Helicoverpa armigera-osaa erityisesti tunnistaa feromoni antagonisti, (Z)-11-hexadecenol (Chang H. et al., 2017). Kirjoittajat kannattivat OR16: n erityisroolia ottaen huomioon, että H., armigera naaraat päästävät antagonisti yhdiste yhdessä sen sukupuoli feromoni ((Z)-11-hexadecenal ja (Z)-9-hexadecenal) strategia välttää ei-optimaalinen pariutumisen kanssa epäkypsä miehillä. Varsinkin kun OR16 oli kaatanut genomin muokkaus tekniikka CRISPR/Cas9 ja H. armigera miehillä testattiin elektrofysiologia ja käyttäytymisen määrityksissä, ei EAG vastaus oli kirjattu ja urokset yritti paritella epäkypsä naisilla.
Hajureseptorit vs., Sitovia Proteiineja: Plussat ja Miinukset Hyönteisten Tuholaisten Hallinta
tapauksessa OBPs, että ligandin spesifisyys ja mekanismeja OBPs edustavat kiistanalaisia näkökohtia, joka näyttää olevan vahvasti riippuvainen menetelmät, joita käytetään mittaus ligandin affiniteetti. Esimerkiksi, se on raportoitu, että PBPs, kuten koit’ P. xylostella ja Eogystia hippophaecolus, voivat sitoa sekä sukupuoli feromoni osia ja analogit (Sun et al., 2013A; Hu ym., 2018). Tämä viittaa siihen, että jatkokäyttäjät, kuten ORs, voisivat parantaa hajuaistin vastaanoton erityisyyttä ja herkkyyttä., Viimeaikaiset todisteet tukevat sitä, että PBPs: n ja PRs: n yhteisilmaisu voi lisätä herkkyyttä feromoneja kohtaan. Esimerkiksi, useita yhdistelmiä PR1-4 ja PBP1-4 käytettiin testata niiden reaktio sukupuoli feromoni osat koi Chilo suppressalis (Chang et al., 2015). Kirjoittajat totesi merkittävä kasvu herkkyys vastausta kohti (Z)-11-hexadecenal kun PR4 ja PR6 olivat co-ilmaistaan PBP4., Vaikka vuorovaikutus näiden proteiinien voi syntyä uuden tutkimuksen tasoa, kuten tuholaistorjunta tavoitteet, eri pariksi PRs ja PBPs irtoa valot monimutkaisuus hajuaistin järjestelmä hyönteisiä, jolloin lähestymistapa on vaikea tehtävä, sillä suuri joukko yhdisteitä ja proteiineja testata. Huolimatta edellä, hyönteisten OBPs ovat pienten molekyylien koko, helppo tuotannon rekombinantti proteiineja, mikä tekee niistä suosikki tavoitteet rakenteellisia tutkimuksia ja nopeasti sitova seulonta., Esimerkiksi ligandin seulonta kanssa OBPs saattaa sallia tunnistaminen kemialliset ominaisuudet paremmin sitova, kuten ketjun pituus, molekyyli-tilavuus, toiminnallisia ryhmiä, ja bond unsaturation. Nämä, yhdistettynä uusien proteiinien rakenteen ennustaminen menetelmiä käytetään suunnittelussa lääkkeet ja vasta-aineet, kuten homologia mallinnus, dynamics simulaatioita, ja molekyyli-telakka, voisi paikka hyönteisten OBPs suosikki asema yli ORs kuten tavoitteet valvonnan kehittäminen aineet tuholaistorjunnassa.
hyönteisten Orit näyttävät olevan tarkemmin viritettyjä odorantoihin kuin Obpit., ORs: n osoittama suurempi spesifisyys ja tiettyjen reseptoreiden aktivointi/inhibitio tietyn käyttäytymisen vuoksi tekevät näistä proteiineista houkuttelevia kohteita tuholaisten käyttäytymisen manipuloimiseksi. Toteutettavuus esto joko ORx/ORco monimutkainen tai ORco, jonka antagonistit koostuu lupaava strategia häiritä hyönteisten erityinen käyttäytyminen, kuten pariutumisen kautta sukupuoli feromoni-reseptoreihin. Rakenteellisen tiedon puute on kuitenkin pullonkaula, kun hyönteistuhoja käytetään semiokemikaalisten aktiivisuusennusteiden kohteina., Taulukoissa 1, 2 yhteenveto määrä OBPs ja ORs, jotka on tunnistettu hyönteinen lajeja transcriptome (eli RNA-seq) ja genomin sekvensointi. Useimmilla tähän mennessä tutkituilla hyönteisillä on perimätutkimusten mukaan vähintään kaksi ORs-arvoa OBP-arvoa enemmän. Lisäksi, on olemassa laaja laajentamiseen ORs sosiaalisten hyönteisten Hymenopteran järjestyksessä, kuten mehiläinen A. mellifera 170 ORs (Weinstock ym. 2006), ja muurahaiset Solenopsis invicta-ja Cerapachys biroi 400 ja 506 ORs, vastaavasti (Wurm ym., 2011; Oxley ym., 2014)., Samoin, TAI laajennus on myös selvää, että jotkut maatalouden tuholaisia, kuten punainen jauhojen beetle T. castaneum kanssa 265 ORs verrattuna 47 OBPs (Richards et al., 2008). Tämä on vaativa tehtävä, tavoite TAI valinta yhdessä vaikeuksia toiminnallinen ilmaisu läpäisevä proteiineja, kuten syrjäisimmät alueet, jotta näytön suuri määrä ligandeja. Taulukossa 3 esitetään yhteenveto tärkeistä ominaisuuksista sekä OBPs-että ORs-järjestelmässä.
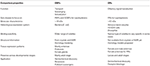
Taulukko 3., Hyönteisten orien ja OBPs: n likimääräinen vertailu ominaisuuksien mukaan.
Lisää Näkökulmia
toiminnallinen luonnehdinta hyönteisten ORs sekä niiden todistettu rooleja hyönteisten hajuaisti on vuodattanut valot herkkyys ja spesifisyys näiden hyönteisten-erityisiä proteiineja. Nämä ennakot on edelleen parantaa niiden toteutettavuus kuten tuholaistorjunta tavoitteet ymmärrystä molekyylitason tunnustamisen mekanismit ja combinatory vuorovaikutus OBPs., Toisaalta, nykyinen massiivinen vaivaa tunnistaminen ja sitova luonnehdinta OBPs useita maatalouden tärkeä hyönteinen lajeja jatkaa ja tarjota enemmän tietoa niiden toimintoja hyönteisten fysiologia. Näin ollen tämän tarkastelun ehdottaa, koska tärkein etu OBPs yli ORs, saatavuus 3D-kide-ja NMR-rakenteita, jotka loppupään lähestymistapoja, kuten homologia mallinnus (tarvittaessa), molekyyli-telakka ja molecular dynamics, olisi tarkentaa hakua bioaktiivisia kemikaaleja., Tämä viime vuonna täydentää ligandin affiniteetti mittaus nopeuttaa tutkimuksen hyönteisten OBPs harkittava uudelleen, koska tavoitteet semiochemical löytö ja työkalut suunnittelu super-ligandien pest control management.
ulkonäkö ja kehitys hyönteismyrkky vastus hyönteisten tuholaisia ovat johtaneet siihen, että intensiivinen tutkimus hyönteisten hajuaisti ja mekanismeja, jotka ovat mukana hermo käsittely. Se on vakiintunut, että useat reseptorit ja entsyymit hyönteisten CNS ovat tavoitteet hyönteisten resistenssin kehittymiseen (Kuva 1)., On osoitettu, että asetyylikoliiniesteraasin (AChE) vuonna liukoisessa muodossa tarjoaa vastustuskykyä orgaaniset fosfori-ja karbamaatti hyönteismyrkyt, toimii bioscavengers (Lee et al., 2015). Samoin, useita hyönteismyrkky vastus mekanismit on osoitettu kirva M. persicae, johon carboxylesterases -, natrium-kanavat, γ-aminovoihappo (GABA) ja nAChR (Bass et al., 2014)., Yhtä tärkeää komponentit hyönteisten reuna hermo järjestelmän ja keskeisten toimijoiden hyönteisten käyttäytymistä, sekä hyönteisten OBPs ja ORs edustavat vaihtoehtoisia tavoitteita tunnistaminen yhdisteiden kanssa semiochemical toimintaa (tai agonistinen vaikutus) ja työkaluja suunnittelun vahva antagonistit parantaa toivottuja käyttäytymisreaktioita hyönteisten tuholaisia ja vähentää hyönteismyrkkyjen käyttöä ja myöhemmin vastus.
Kirjoittaja Maksut
HM kirjoitti kohdat noin OBP: n rakenne ja ORs, kehitetty taulukot ja kuva., J-JZ suunniteltu idea tarkistaa artikla, kirjoitti tärkein kohta, kuten käyttöönotto, OBPs toiminta ja rakenne sekä TAI-OBP vertailu.
eturistiriita Lausunto
kirjoittajat ilmoittavat, että tutkimus on tehty ilman mitään kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdollisia eturistiriitoja.
Kiitokset
tekijät haluavat kiittää FONDECYT 3170433., J-JZ on kiitollinen Koillis-Normaaliyliopiston ja Kiinan Jilinin yliopiston taloudellisesta tuesta sapattivapaalleen opiskella Kiinassa. Rothamsted Research saa apurahan, johon on lisätty rahoitusta bioteknologia ja biologiset tieteet (BBSRC), Iso-Britannia.
Gonzalez, F., Witzgall, P., ja Walker, W. B. (2017). Antennal transcriptomes kolme tortricid koit paljastaa oletetun säilytetty chemosensory reseptoreihin sosiaali-ja elinympäristö hajuaistin vihjeitä. Sci. Torjunnat 7, 1-12. doi: 10.,1038/srep41829
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar
Kaissling, K. E. (2013). Kinetiikka hajuaistin vastauksia saattaa paljolti riippua tuoksu-reseptorin vuorovaikutus ja tuoksu deaktivointi postulated for flux ilmaisimet. J. Comp. Fysiol. Neuroetoli. Aistihermo Käyttäytyy. Fysiol. 199, 879–196. doi: 10.1007/s00359-013-0812-z
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar