Tervezése, építése, randomizált könyvtárak assaying Cas9 PAM beállítások
PAM könyvtárak tartalmazó randomizált DNS-szekvenciák azonnal downstream a DNS-szekvencia, kiegészítő távtartó útmutató RNS keletkezett használt empirikusan meghatározni a PAM elismerés a II-es Típusú Cas9 endonucleases (Fig. 1)., A vezető RNS távtartó célszekvenciájának rögzítésével a randomizált bázisok szubsztrátként szolgálnak a Cas9 endonukleáz Pam specifikusságának közvetlen kiolvasásához. Randomizált szekvenciákat vezettek be egy plazmid DNS vektor A Pam régióban egy protospacer cél szekvencia bizonyítja tökéletes homológia a vezető RNS távtartó T1 (CGCUAAAGAGGAAGAGGACA). Két könyvtár mérete és összetettsége öt randomizált bázispárból (1,024 potenciális PAM kombináció) hét randomizált bázispárra (16,384 potenciális PAM kombináció) nőtt., Az 5 bp könyvtár randomizációját egyetlen öt véletlenszerű maradékot tartalmazó oligonukleotid szintézisével vezették be. Az egyszálú oligonukleotidot a PCR (1. kiegészítő fájl: S1A ábra) kettős szálú sablonná alakította át, a plazmidvektorba klónozva (1.kiegészítő fájl: S1B Ábra), és E. coli-ba átalakítva, a módszerek szakaszban leírtak szerint., A 7 bp Pam könyvtár optimális véletlenszerűségének biztosítása érdekében a könyvtár méretét és összetettségét úgy csökkentettük, hogy négy oligonukleotidot szintetizáltunk, amelyek mindegyike hat véletlenszerű maradékot tartalmaz, valamint egy hetedik rögzített maradékot, amely G, C, A vagy T-t tartalmaz. A négy oligonukleotid mindegyike külön-külön átalakult kettős szálú DNS-be, amelyet a módszerek szakaszban leírtak szerint pTZ57R/T vektorrá klónoztak, és az 5 bp könyvtár leírásai szerint E. coli-ba transzformáltak., Miután átalakulás, plazmid DNS-t találtak, valamint kombinált mind a négy 6 bp PAM könyvtárak, hogy létrehoz egy randomizált 7 bp PAM könyvtár, amely 16,384 lehetséges PAM kombinációk. Mindkét könyvtár esetében a véletlenszerűség beépülését mély szekvenálással validálták; a nukleotidösszetétel vizsgálata a Pam régió minden helyzetében egy pozíciófrekvencia-mátrix (PFM) segítségével (metódusok szakasz és ) (további 1. fájl: S2A és B ábra)., Az 5 bp-es és 7 bp-es randomizált Pam-könyvtárban az egyes PAM-szekvenciák eloszlását és gyakoriságát további 1-es fájlban mutatjuk be: S3 és S4 számok.
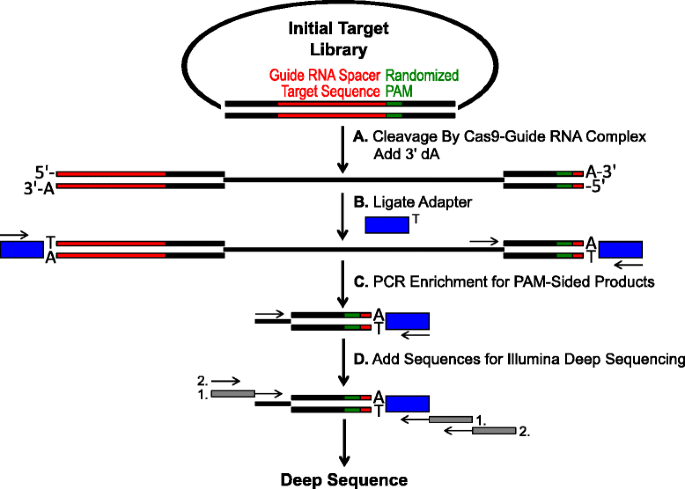
sematikus azonosítására Pam preferenciák Cas9 hasítás in vitro. a kezdeti plazmid könyvtár randomizált PAM (zöld doboz) hasítjuk Cas9 komplex 3 ” dA túlnyúlások adunk. b Adapterek 3 ‘ dT túlnyúlás (kék doboz) vannak kötve mindkét végén a hasítás termék., a C alapozókat a PAM-oldalú hasított termékek PCR általi gazdagítására használják., d Után PCR gazdagodás, a DNS fragmentumokat tisztított, valamint Illumina kompatibilis rögzíti, vonalkódok vagy ‘farkú-on keresztül két kör PCR (szürke dobozok) pedig Illumina mély szekvenált
Assaying Cas9 PAM beállítások
A randomizált PAM könyvtárak az előző szakaszban leírt voltak kitéve in vitro emésztés különböző koncentrációban rekombináns Cas9 fehérje előre útmutató RNS annak érdekében, hogy a vizsgálat Cas9 endonuclease PAM beállítások a dózis-függő módon., Lebontás után a Cas9-útmutató RNS ribonucleoprotein (RNP) komplexek, PAM sorozat kombinációk a randomizált PAM könyvtár támogatott dekoltázs elfogták vágás adapterek, hogy a szabad végét a plazmid DNS-molekulák hasított a Cas9-útmutató RNS komplex (Fig. 1a és b). A hasított végek hatékony ligálásának és rögzítésének elősegítése érdekében a Cas9 endonukleázok által generált tompa végű kettős szálú DNS-vágást úgy módosították, hogy 3 ‘dA túlnyúlást tartalmazzon, az adaptereket pedig úgy módosították, hogy tartalmazzanak egy kiegészítő 3’ dT túlnyúlást., Termelnek elegendő mennyiségű DNS szekvenálás, DNS-fragmentumok menedéket a PAM sorozat támogatja a dekoltázs volt PCR segítségével felerősített egy alapozó az adapter, a másik közvetlenül szomszédos, hogy a PAM régió (Fig. 1c). A kapott PCR amplifikált Cas9 Pam könyvtárakat ampli-seq sablonokká alakították át (ábra. 1d) és az amplicon adapteroldaláról mélyreolvasás., A megfelelő lefedettség biztosítása érdekében a Cas9 Pam könyvtárakat legalább ötször nagyobb mélységre szekvenálták, mint a kezdeti randomizált Pam könyvtár sokféleségét (5,120 és 81,920 olvasás az 5 és 7 bp Pam randomizált könyvtárak esetében). PAM sorozatok azonosítottak a kapott szekvencia adatok csak kiválasztja azokat olvassa, amely egy tökéletes 12 nt sorozat mérkőzés kísérő mindkét oldalán, az 5-ös vagy 7 nt PAM sorozat (attól függően, hogy a randomizált PAM könyvtár használt); elfog csak azok a PAM sorozatok eredő tökéletes Cas9-útmutató RNS célterületre elismerés, valamint a dekoltázs., A kezdeti randomizált PAM könyvtárakban rejlő torzítás kompenzálására az egyes PAM-szekvenciák frekvenciáját a kezdőkönyvtár frekvenciájára normalizálták. Mivel az itt leírt vizsgálat közvetlenül rögzíti a Cas9 hasítható Pam-szekvenciákat, valószínűségi modellezést alkalmaztak az egyes Cas9-fehérjék PAM-konszenzusának kiszámításához. Ez volt elérni értékeli a valószínűsége, hogy megtaláljuk az egyes nukleotid (G, C, A, vagy T) minden helyzetben a PAM sorozat önállóan használja a helyzetben, frekvencia mátrix (PFM) (Módszerek részben )., A kapott valószínűségeket ezután WebLogo-ként vizualizálták .
a hamis pozitívok hajlandóságának vizsgálatához a Cas9 RNP komplexek hozzáadását az emésztési lépésben kihagyták (ábra. 1a) a vizsgálatot a PCR dúsítási lépésen keresztül végezték (ábra. 1c). Amint az az 1. kiegészítő fájlban látható: az S5A ábra, a Cas9-vezető RNS komplexek hiányában nem észleltek amplifikációs termékeket. Így, jelezve, hogy a hamis pozitívumok előfordulása alacsony, és nem járul hozzá jelentősen a vizsgálat eredményeihez.,
A Streptococcus pyogenes és a Streptococcus thermophilus (CRISPR3 és CRISPR1 systems) Cas9 fehérjék
a vizsgálat érvényesítése érdekében megvizsgálták a Streptococcus pyogenes (Spy) és a Streptococcus thermophilus CRISPR3 (Sth3) Cas9 fehérjék Pam-preferenciáit, amelyek PAM-szekvencia-követelményét korábban jelentették. Az 5 bp randomizált PAM könyvtárból 1 µg (5,6 nM) in vitro emésztést végeztünk két koncentrációban, 0,5 és 50 nM-en, előre összeszerelt kém-vagy Sth3 Cas9 protein, crRNA és tracrna RNP komplexekkel 1 órán keresztül, 100 µL reakció térfogatban., Az 5 bp-es randomizált PAM könyvtár frekvenciája alapján a Spy és az Sth3 Cas9 Pam szekvenciák (NGG, illetve NGGNG) végső koncentrációja 0,40 nM, illetve 0,11 nM volt az emésztésben. A hasítást alátámasztó Pam-szekvenciákat tartalmazó randomizált Pam-könyvtár tagjai az előző részben leírtak szerint kerültek rögzítésre és azonosításra. Negatív kontrollként a kezdő, tisztítatlan, randomizált PAM könyvtárat szekvenálásnak és PFM elemzésnek vetették alá a Cas9 RNP komplexeknek kitett könyvtárak mellett., Amint az az 1. kiegészítő fájlban látható: S5B és C ábra, Cas9 RNP komplex emésztés hiányában nem léteznek szekvencia-preferenciák, amint azt az egyes nukleotidok közel tökéletes eloszlása mutatja A PAM minden pozíciójában a PFM táblázatban, valamint a WebLogo informatív tartalmának hiánya az ellenőrzéshez. Ez a stark constrast füge. 2a és b, amely a kém és Sth3 Cas9 RNP komplexekkel megemésztett könyvtárakból származó szekvenciák összetételét szemlélteti. A PFM-ből származó WebLogos vizsgálata (ábra., 2a és b) A kém, illetve az Sth3 Cas9 fehérjék kanonikus PAMPREFERENCIÁINAK jelenlétét is feltárják , NGG és NGGNG. Bár a Spy és az Sth3 Cas9 fehérjékre vonatkozó PAM-preferenciákat mind a 0,5 nM-es, mind az 50 nM-es digestekben megfigyelték,a specificitás általános kiterjesztése az 50 nM-es digest körülmények között. Ez a legnyilvánvalóbb a Spy Cas9 fehérje 2. pozíciójában, ahol a nem kanonikus a maradék gyakorisága drámaian megnő (ábra. 2a)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
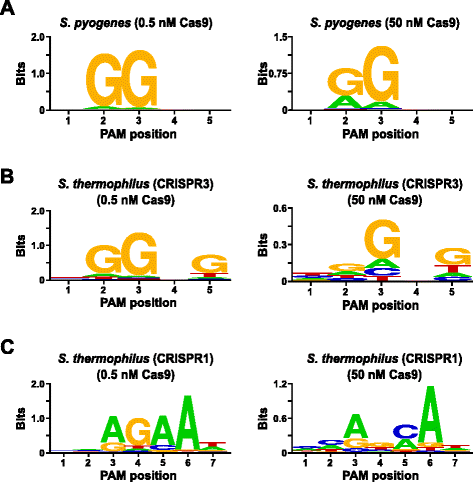
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., Gyakorisága nukleotid minden PAM helyzetben volt, függetlenül kiszámítása a helyzetben, frekvencia mátrix (PFM), illetve ábrázolni, mint egy WebLogo
További érvényesítése a vizsgálatot végezte vizsgálja a PAM beállítások a Streptococcus thermophilus CRISPR1 (Sth1) Cas9 fehérje, amelynek PAM sajátossága beszámoltak arról, hogy meghosszabbítja ki, hogy 7 bp . A 7 bp randomizált PAM könyvtár 1 µg (5,6 nM)-ját sablonként használva az Sth1 Cas9-vezető RNS-emésztést két koncentrációban, 0,5 nM-en és 50 nM-en, az RNP komplexben végeztük el a fent leírtak szerint., Kontrollként Spy és Sth3 Cas9 RNP komplexeket is használtak a 7 bp randomizált Pam könyvtár megemésztésére, de csak egyetlen, 0,5 nM-es, RNP komplex koncentrációban. A 7 bp randomizált Pam könyvtár frekvenciája alapján a korábban jelentett Pam szekvenciák az Sth1 (NNAGAAW), a Spy (NGG) és az Sth3 (NGGNG) esetében 0,01 nM, 0,22 nM, illetve 0,05 nM végső koncentrációban voltak., Amint azt az 1. kiegészítő fájl is mutatja: az S6A és B ábra, a 7 bp könyvtár segítségével előállított Spy és Sth3 Cas9 fehérjék PAM-preferenciái majdnem azonosak voltak az 5 bp könyvtárral, amely erős bizonyítékot szolgáltat a vizsgálat reprodukálhatóságára. A Pam preferenciák az Sth1 Cas9 fehérje is szorosan illeszkedik, hogy a korábban bejelentett, NNAGAAW, a 0,5 nM Cas9-guide RNS komplex koncentráció (ábra. 2c)., Hasonló Kém, valamint Sth3 Cas9 fehérjék, Sth1 Cas9 volt képes hasító egy sokszínűbb készlet PAM szekvenciák az a reakció, amely egy magasabb koncentrációja Cas9-útmutató RNS komplex (50 nM), a legszembetűnőbb az volt, hogy a jelölt veszteség a G maradék követelmény a helyzetben, 4 pedig a közel egyenlő inkább a C, bp pozíció 5 (Fig. 2c). Ez eltérő PAM konszenzust eredményezett, mint az alacsonyabb koncentrációkban.,
annak megvizsgálására, hogy a Pam specificitása független-e a vezető RNS típusától , a kettős crrna:tracrRNA vagy sgRNA, Spy, Sth3 és Sth1 Cas9 Pam preferenciákat bináris Cas9 és sgRNA RNP komplex segítségével is megvizsgálták. Az emésztést Egyetlen 0,5 nM-es RNP komplex koncentrációban végezték, a PAM preferencia analízist pedig a fent leírtak szerint. Amint az az 1. kiegészítő fájlban is látható: S7A, B és C ábra, a PAM preferenciái közel azonosak voltak, függetlenül az alkalmazott vezető RNS típusától; vagy egy crRNA: tracrna duplex vagy sgRNA., Ezen túlmenően annak megerősítése érdekében, hogy a Pam specificitását nem befolyásolja nagymértékben a cél DNS vagy távtartó szekvencia összetétele, az 5 vagy 7 bp randomizált könyvtár másik oldalán lévő szekvenciát egy másik távtartóval történő hasításra irányították; T2-5 (UCUAGAUAGAUUACGAAUUC) az 5 bp könyvtárhoz vagy T2-7 (CCGGCGACGUUGGGUCAACU) a 7 bp könyvtárhoz. Spy és Sth3 Cas9 fehérjék előtelepített sgRNAs célzó T2 szekvenciát használtunk kihallgatni a 5 bp randomizált Pam könyvtár, míg az Sth1 Cas9-T2 sgrna komplexek használták megemészteni a 7 bp randomizált Pam könyvtár., A PAM preferenciáit a fent leírtak szerint határozták meg. A Pam preferenciák mind a 3 Cas9 fehérje esetében közel azonosak voltak, függetlenül a távtartótól és a cél DNS-szekvenciától (további 1.fájl: S8A, B, és C ábra).
az sgRNA és a Pam preferenciák azonosítása a Brevibacillus laterosporus Cas9 fehérjére
annak érdekében, hogy empirikusan megvizsgálják a Pam preferenciáit egy olyan Cas9 fehérjére, amelynek PAM-ját nem határozták meg, a Brevibacillus laterosporus ssp360d4 (Blat) törzsből származó II-C típusú CRISPR-Cas Locust a Cas9 ortológokra vonatkozó belső DuPont Pioneer adatbázisok keresésével azonosították., A locus (körülbelül 4,5 kb) egy cas9 gént tartalmazott, amely képes egy 1,092 polipeptid kódolására, egy CRISPR tömböt, amely hét ismétlődő távtartó egységet tartalmaz közvetlenül a cas9 gén után, és egy tracrRNA kódolási régiót, amely a cas9 gén előtt helyezkedik el, részleges homológiával a CRISPR tömb ismétléséhez (ábra. 3a). Az ismétlés és a távtartó hossza (ennek megfelelően 36 és 30 bp) hasonló más II. típusú CRISPR-Cas rendszerekhez, a nyolc ismétlés közül öt 1 vagy 2 bp mutációt tartalmaz (1.ábra). 3b és további 1. fájl: S9 ábra)., Más gének, amelyek jellemzően egy II. típusú CRISPR-Cas Locusban találhatók, vagy csonkoltak (cas1) vagy hiányosak voltak (ábra. 3a).
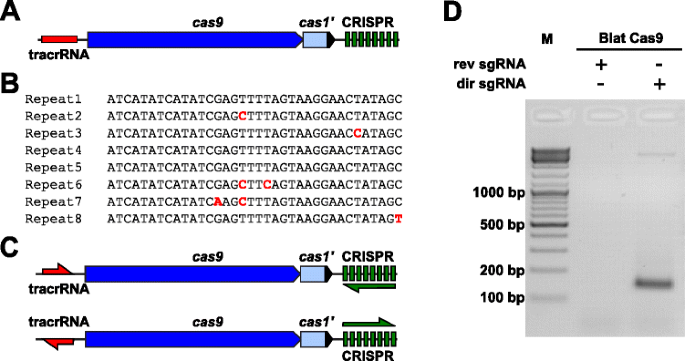
II típusú CRISPR-Cas elemek azonosítása a Brevibacillus laterosporus SSP360D4 CRISPR-Cas rendszerben. a brevibacillus laterosporus SSP360D4 II. típusú CRISPR-Cas rendszer genomikus DNS-régiójának szemléltetése. b A Brevibacillus laterosporus SSP360D4-ben azonosított II. típusú CRISPR tömb ismételt szekvenciák összehasonlítása., c a BREVIBACILLUS laterosporus SSP360D4 II. típusú CRISPR-Cas rendszerre vonatkozó “közvetlen” és “fordított” tracrrns és CRISPR array transzkripciós forgatókönyvek. d agarózgél reakciótermékekkel, jelezve, hogy csak a”közvetlen”sgRNA (dir sgRNA), de nem a “fordított” sgRNA (rev sgRNA) támogatja a plazmid könyvtár hasítását a Brevibacillus laterosporus SSP360D4
az útmutató RNS-követelmény a blat Cas9 fehérjét két sgrna variáns előállításával határoztuk meg., Ezeket a változatokat úgy hozták létre, hogy figyelembe vegyék mind a tracrRNA, mind a CRISPR tömb lehetséges érzékelési vagy érzékellenes expressziós forgatókönyveit (ábra. 3c), és arra használták, hogy a randomizált Pam könyvtárban megvizsgálják, melyik kifejezési forgatókönyv támogatta a blat Cas9 hasítási aktivitását. Az egyetlen vezető RNS-t úgy tervezték meg, hogy először azonosítják a feltételezett tracrRNA molekulák határait azáltal, hogy elemzik azokat a régiókat, amelyek részben kiegészítik az ismétlés 22 nt 5 ” terminusát (anti-repeat)., Ezután a tracrRNA 3′ végének meghatározásához lehetséges másodlagos struktúrákat és Terminátorokat használtunk a downstream fragmentum végződési régiójának előrejelzésére. Ezt úgy hajtottuk végre, hogy a Rho független-szerű végződési szekvenciák jelenlétét szűrtük a Karvelis et al. a környező DNS-t RNS-szekvenciává alakítja, majd a kapott struktúrákat UNAfold segítségével vizsgálja ., A kapott sgRNAs-t úgy tervezték, hogy T7 polimeráz transzkripciós iniciációs jelet tartalmazzon az 5′ végén, majd egy 20 nt célfelismerési szekvenciát, 16 nt crRNA ismétlést, 4 nt önhajtogató hajtűhurkot, valamint a crRNA ismétlési régióját kiegészítő anti-repeat szekvenciát, amelyet a putatív tracrRNA fennmaradó 3′ része követ. Az sgrna változat, amely egy feltételezett tracrrns-t tartalmaz, a cas9 génnel azonos irányba átírva (ábra. 3c) “közvetlen” sgRNA, míg a tracrRNA-t tartalmazó sgRNA ellentétes irányban a “fordított” sgrna., A Blat Cas9 Sgrna RNP komplex ötven nM-ét a 7 bp-es randomizált Pam könyvtár 1 µg (5,6 nM) – jával inkubáltuk. A könyvtári emésztés és a 3′ dA túlnyúlások hozzáadása után az adaptereket ligálták, a dekoltázs termékeket pedig PCR-re erősítették (1.ábra). 1). A reakciótermékek agarózgél-elektroforézissel történő elemzése azt mutatta, hogy a “közvetlen” sgRNA, de nem a “fordított” sgrna támogatta a plazmid könyvtár hasítását (ábra. 3d). A “közvetlen” sgRNA szekvenciáját és várható másodlagos szerkezetét az 1. kiegészítő fájlban, az S10 ábrán mutatjuk be.,
a Blat Cas9-re vonatkozó megfelelő vezető RNS meghatározása után a Pam azonosítást a kém, az Sth3 és az Sth1 Cas9 fehérjék esetében a 7 bp-es randomizált PAM könyvtárhoz hasonlóan végezték el, az előre összeállított Blat Cas9 “direct” Sgrna RNP komplex két koncentrációjával, 0,5 és 50 nM-rel. Amint az ábrán látható. 4a, a PFM WebLogo Pam konszenzus a blat Cas9 fehérje esetében a 0,5 nM-es digest körülmények között NNNNCND (N = G, C, A, vagy T; D = A, G, vagy T) volt, erősen előnyben részesítve a C-t a Pam szekvencia 5.pozíciójában., A PFM-táblázat alapos vizsgálata során a 7.pozícióban mérsékelt preferenciát figyeltek meg az A-ra, a 4. és a G, C, vagy a 6. pozícióban a T-re, illetve a T-re vonatkozóan pedig enyhe preferenciákat figyeltek meg (1. kiegészítő fájl: S11. ábra). A Spy, az Sth3 és az Sth1 Cas9 fehérjékhez hasonlóan a Pam specificitása is kiszélesedik, mivel a Cas9-sgRNA komplex koncentrációja nő. Ez a legnyilvánvalóbb az 5. helyen, ahol a Pam-szekvenciák nagyobb része, amely 50 nM-en szermaradék-alátámasztó hasítást tartalmaz, összehasonlítva a 0,5 nM-es emésztési feltételekkel.
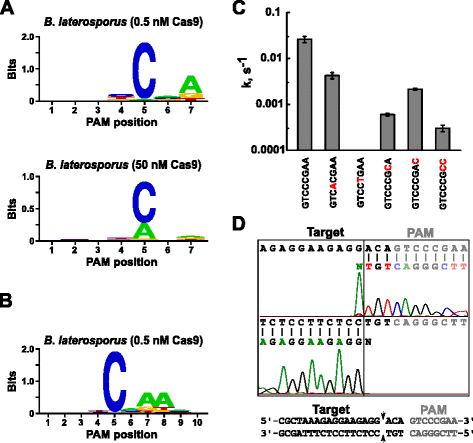
p > a Brevibacillus laterosporus SSP360D4 (Blat) Cas9 enzim dekoltázspozíciói. Blat Cas9 PAM preferenciák amikor 1 µg könyvtár DNS-t hasítottak 0,5 nM vagy 50 nM Cas9-sgRNA komplex (a), kiterjesztették a 10.pozícióra azáltal, hogy a protospacer célt 3 bp (b) – rel eltolták. A nukleotidok gyakoriságát minden egyes Pam-pozícióban egy pozíciófrekvencia-mátrix (PFM) segítségével önállóan számították ki, és Weblogóként ábrázolták ., c A gtcccgaa Pam szekvenciában mutációkat tartalmazó (piros színnel jelzett) szuperkoilált plazmid DNS-szubsztrátok hasítási aránya. Minden adatpont a ≥3 független kísérlet átlagos értéke. Hiba sávok kapnak, mint a S. D. d Run-off sorozatot, hogy mindkét értelemben pedig anti-értelemben irányban plazmid DNS-hasított a Blat Cas9
Mivel Blat Cas9 elfogadhatja, minden alap az első három pozíció a PAM sorozat (Fig. 4a), a T1 távtartót három nukleotid eltolta az 5 ” irányba, hogy lehetővé tegye a PAM azonosítását 7-ről 10 bp-re., Az eltolódott T1 távtartót, a T1-3-at (aaacgcuaaagaggaagagagagg) beépítették a Blat “direct” sgRNA-ba, és a PAM azonosítást a Spy, Sth3, Sth1 és Blat Cas9 fehérjék esetében korábban leírtak szerint végezték el. PAM preferencia elemzés feltárta a Pam specificitása Blat Cas9 lehet terjeszteni ki helyzetben 8 ahol van egy mérsékelt előnyben egy további A (ábra . 4b).
Pam specificitását a blat Cas9 esetében plazmidok generálásával igazolták, hogy mutációkat tartalmazzanak a Pam legtartósabb maradékaiban (ábra. 4c)., A C nukleotid 5. pozícióban történő cseréje megszüntette a plazmid DNS hasítását, megerősítve annak kulcsszerepét a Blat Cas9 Pam felismerésében. A nukleotidok 7.és 8. pozícióban történő helyettesítése jelentősen csökkent (43×, illetve 12×) a túlhűtött plazmid hasadási sebessége szintén jelzi ezeknek a nukleotidoknak a fontosságát a Blat Cas9 Pam felismerésben.,
azonosítása a cél DNS hasítás pozíciók a Blat Cas9 fehérje, egy plazmid, amely 20 bp régió megfelelő a távtartó T1 követi PAM sorozat, GTCCCGAA, esik a PAM konszenzus a Blat Cas9, NNNNCNDD, keletkezett, s emésztett a Blat Cas9-útmutató RNS ribonucleoprotein összetett. Közvetlen DNS-szekvenálást alkalmaztak a blat Cas9 RNP komplex által generált lineáris DNS-molekula végeinek meghatározására. A szekvencia eredményei megerősítették, hogy a plazmid DNS hasítása a Pam szekvencia protospacer 3 nt 5′ – jében történt (ábra., 4D) hasonló a kém, Sth3 és Sth1 Cas9 fehérjék esetében megfigyelthez .
A planta Genomszerkesztésben a blat Cas9 és az sgRNA
használatával az Sgrna és a Pam preferenciáinak tisztázását követően a kukorica optimalizált Cas9 és sgRNA expressziós kazettákat hoztak létre a planta vizsgálat során, amint azt korábban az S. pyogenes cas9 gén és az sgRNA esetében leírták . Röviden, a blat cas9 gént kukorica kodonra optimalizálták, a burgonya ST-LSI gén intron 2-jét pedig beillesztették az E. coli expressziójának megzavarására, és megkönnyítették a planta optimális összekapcsolását (további 1.fájl: S12 ábra)., A Blat Cas9 fehérje nukleáris lokalizációját a kukoricasejtekben megkönnyítette mind az amino, mind a karboxil-terminális nukleáris helyek jeleinek, SV40 (MAPKKRKV), valamint az Agrobacterium tumefaciens VirD2 (KRPRDRHDGELGGRKRAR) hozzáadása (További fájl 1: S12 ábra). A Blat cas9 gént a növényi sejtekben úgy fejezték ki, hogy az optimalizált cas9-et egy kukorica Ubiquitin promoterhez és pinII terminátorhoz kapcsolták plazmid DNS vektorban., Előnyt jelent hatékony sgRNA kifejezés, a kukorica, a sejtek, a kukorica U6 polimeráz III szervező, illetve a terminátor (TTTTTTTT) volt elszigetelt, olvasztott, hogy az 5′ 3′ végén egy módosított Blat sgRNA kódoló DNS-szekvencia, illetve (Előfordulhat, hogy a fájl 1: Ábra S13). A módosított Blat sgRNA az in vitro vizsgálatokban alkalmazott két módosítást tartalmazott; a T-G változás a 99. pozícióban és a T-C módosítás az sgRNA 157. pozíciójában (további 1.fájl: S13 ábra). A változásokat azért vezették be, hogy eltávolítsák a potenciális korai U6 polimeráz III végződési jeleket a Blat sgRNA-ban., Az in vitro vizsgálatokban alkalmazott változathoz képest az sgrna másodlagos szerkezetére minimális hatást gyakorló változások (az adatok nem voltak kimutatva).
a spy és Blat Cas9 hasításából eredő DNS kettős szálú törések (DSBS) tökéletlen, nem homológ végcsatlakozásából (NHEJ) eredő mutációs hatékonyság pontos összehasonlításához a protospacer azonos genomikus célhelyeket úgy választották ki, hogy a spy és Blat Cas9 kompatibilis PAMs, NGGNCNDDD célokat azonosítottak., A Blat és a Spy Cas9 esetében azonos távtartó szekvenciákat választottunk ki úgy, hogy a 18-21 nt szekvenciát közvetlenül a PAM előtt rögzítettük. Annak érdekében, hogy az optimális U6 polimeráz III expresszió ne vezessen be eltérést az sgRNA távtartón belül, az összes célszekvenciát úgy választottuk ki, hogy az 5 ” végén természetesen G-ben megszűnjön. A célértékeket az Ms45 kukorica termékenységi gén 1.és 4. exonjában, valamint a kukorica liguleless-1 géntel szemben lévő régióban határozták meg és választották ki.,
a blat Cas9 kukoricában történő mutációs aktivitását biolisztikusan átalakították 10 napos éretlen kukorica embriók (IMEs) cas9 és sgRNA géneket tartalmazó DNS-vektorokkal. A Blat és az azzal egyenértékű Spy Cas9 és sgRNA expressziós vektorokat egymástól függetlenül vezették be a kukorica Hi-Type II IMEs-be a benne leírtakhoz hasonló részecskefegyver-transzformációval . Mivel a részecske fegyvert átalakulás lehet nagyon változó, vizuális DNS marker kifejezés kazetta, Ds-Vörös, volt is co-szállított a Cas9, valamint sgRNA vivővektorok, hogy a támogatást a kiválasztott egyenletesen átalakult Írásjegybevivő., Összesen három átalakulás ismétlések végezték 60-90 Írásjegybevivő pedig 20-30 a legtöbb egyenletesen átalakult Írásjegybevivő az egyes párhuzamos betakarították 3 nap után átalakulás. Teljes genomiális DNS-t kivonták a környéket a cél helyén volt erősítik PCR, valamint amplicons szekvenált, hogy egy olvasni mélység meghaladja a 300 000-et. Az így kapott leolvasásokat megvizsgáltuk mutációk jelenlétére a hasítás várható helyén, összehasonlítva a kontroll kísérletekkel, ahol az sgrna DNS expressziós kazettát kihagyták az átalakulásból. Amint az ábrán látható., 5A, mutációkat figyeltek meg a blat Cas9 hasításának várható helyén, a leggyakoribb mutációk az egyetlen bázispár beillesztése vagy törlése. Hasonló javítási mintákat figyeltek meg a Spy Cas9 protein esetében is (további 1. fájl: S14 ábra). A blat Cas9 mutációs aktivitása a vizsgált három helyszín közül kettőnél robusztus volt, és megközelítőleg 30% – kal haladta meg az Ms45 exon 4 célhelyen lévő Spy Cas9-ét (ábra). 5b).
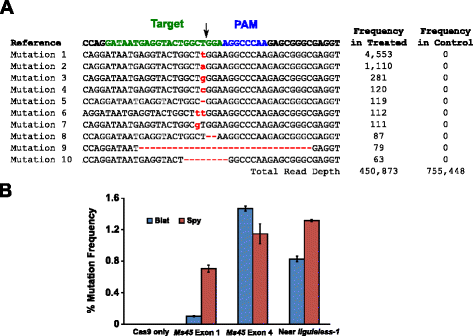
Brevibacillus laterosporus Cas9 elősegíti az NHEJ mutációkat a kukoricában. az Ms45 gén exon 4-ben a blat Cas9-ben észlelt NHEJ mutációk Top 10 legelterjedtebb típusa. A fekete nyíl jelzi a hasítás várható helyét; a mutációk piros színnel vannak kiemelve; az alsó betűkészlet beillesztést jelez; ” – ” törlést jelez. b A Spy és a Blat Cas9 NHEJ mutációs gyakoriságának összehasonlítása a kukorica három protospacer azonos célhelyén. Az NHEJ mutációkat az átalakulás után 3 nappal mély szekvenálással detektálták., Hiba rudak képviselik SEM, n = 3 részecske Pisztoly transzformációk. A Cas9 csak a negatív kontroll, és a PCR amplifikációból és szekvenálásból eredő mutációk átlagos (mindhárom célhelyen) háttérfrekvenciáját jelenti