anche se i composti volatili con attività agonista sono stati proiettati contro ORs, una specifica sostanza chimica con un forte agonista effetto sulle zanzare, 2-(4-Etil-5-(pyridin-3-yl)-4H-1,2,4-triazol-3-ylthio)-N-(4-etilfenil)acetammide (VUAA1), ha aperto il campo di ricerca (Jones et al., 2011; Taylor et al., 2012). Studi successivi di Taylor et al., (2012) ha fornito prove di sostanze chimiche derivate da VUAA1, come VUAA4, in grado di aumentare il suo effetto agonista di 10 volte su ORco da A. gambiae, H. virescens e Harpegnathos saltator. È interessante notare che gli autori hanno riferito che qualsiasi cambiamento sui sostituenti ammidici causerà una completa perdita di attività agonista. Ciò fornisce informazioni utili sui requisiti strutturali degli agonisti e sulla relazione struttura-attività tra analoghi VUAA e RUP. Infine, nonostante l’attività agonista migliorata dei prodotti chimici VUAA, il suo peso molecolare relativamente alto (367,47 g mol−1 per VUAA1) vs., agonisti volatili, come bombykol (238.42 g mol−1), rende una consegna volatile diretta di VUAA qualcosa non fattibile. Con questo in mente, la ricerca di analoghi strutturali più piccoli rappresenta un interessante focus di ricerca.
Antagonismo sulla subunità ORco
Insieme allo studio di analoghi legati alla VUAA che possono agire come forti agonisti, è emerso anche il blocco di ORco da parte degli antagonisti per guidare la progettazione di semiochimici e pesticidi., Pertanto, un analogo strutturale di VUAA1, VU0183254 (2-(4-Etil-5-furan-2-il-4H-triazol-3-ilsulfanil)-1-fenotiazin-10-il-etanone), è stato segnalato per inibire la risposta di ORco, agendo come modulatore allosterico in A. gambiae e interrompendo il riconoscimento di agonisti come eugenolo dal complesso OR65/ORco (Jones et al., 2012). Altri analoghi VUAA-strutturali sono stati segnalati anche come antagonisti. Un esempio è la N -, 2-sostituito triazolothioacetamide composti OLC3 e OLC12 che interrompe la risposta ORco in modo simile in C. quinquefasciatus, A. gambiae, D. melanogaster, e O., nubilalis, suggerendo un sito di legame conservato in ORco (Chen e Luetje, 2012). Considerando l’inibizione dell’ORco come una strategia promettente per interrompere i comportamenti degli insetti, sembra che gli sforzi successivi dovrebbero mirare ai composti con un peso molecolare inferiore rispetto agli antagonisti derivati da VUAA. Ad esempio, OX1a (232 g mol−1), triptamina (160,22 g mol−1) e isopropil cinnamato (190,24 g mol−1) sono stati segnalati per avere un effetto antagonista su ORco (Chen e Luetje, 2013, 2014; Tsitoura et al., 2015) con circa la metà o meno peso molecolare di VUAA1., Tuttavia, l’uso futuro di questi antagonisti dovrebbe essere studiato attentamente, poiché il blocco dell’ORco conservato può influenzare non solo gli insetti dannosi, ma anche quelli benefici.
Oltre all’effetto antagonista sondato in vitro, l’evidenza a livello comportamentale supporta l’idea che gli analoghi strutturali dei feromoni possano funzionare come antagonisti. Ad esempio, Sellanes et al., (2010) ha riportato l’inibizione della risposta sessuale nella falena della melata Cryptoblabes gnidiella quando gli analoghi strutturali, (Z)-9-tetradecenil formiato e (Z)-11-esadecenil formiato, sono stati aggiunti al feromone sessuale sintetico, (Z)-11-esadecenale e (Z)-13-ottadecenale, nei test in galleria del vento. Questo effetto dell’antagonista del feromone più successivamente è stato corroborato nelle analisi del campo, in cui la cattura dei maschi di C. gnidiella è diminuita in un modello dose-dipendente. L’antagonismo dei feromoni è stato riportato anche per B. mandarina, un antenato di B. mori (Daimon et al., 2012)., Le loro scoperte confermano bombykol come feromone sessuale e bombykal e bombykyl acetato come antagonisti, che hanno fortemente inibito l’attrazione dei maschi in campo al feromone sessuale bombykol. Più recentemente, la prova dell’antagonismo del feromone è stata riferita per la falena del muso Herpetogramma submarginale. Quando (Z)-13-esadecenolo è stato aggiunto al suo feromone sessuale, (Z) – 13-esadecenil acetato, ha diminuito significativamente il numero di maschi catturati in campo (Yan et al., 2015)., L’antagonismo del feromone sembra basato sulle differenze nel gruppo funzionale chimico quali gli alcoli, le aldeidi e gli esteri secondo la specie dell’insetto. Tuttavia, l’effetto antagonista di questi analoghi strutturali potrebbe non essere dovuto all’inibizione di ORco ma alla specificità di ORx rispetto agli antagonisti. Uno studio recente suggerisce che l’OR16 di Helicoverpa armigera è in grado di riconoscere specificamente l’antagonista del feromone, (Z)-11-esadecenolo (Chang H. et al., 2017). Gli autori hanno sostenuto il ruolo specifico di OR16 considerando che H., le femmine di armigera emettono il composto antagonista insieme al suo feromone sessuale ((Z)-11-esadecenale e (Z)-9-esadecenale) come strategia per evitare l’accoppiamento non ottimale con i maschi immaturi. In modo eccezionale, quando l’OR16 è stato abbattuto dalla tecnica di editing del genoma CRISPR / Cas9 e H. armigera maschi sono stati testati mediante elettrofisiologia e saggi comportamentali, nessuna risposta EAG è stata registrata e maschi hanno cercato di accoppiarsi con femmine immature.
Recettori odoranti vs., Proteine leganti: pro e contro per la gestione dei parassiti degli insetti
Per il caso di OBPs, la specificità del ligando e i meccanismi di OBPs rappresentano aspetti controversi, che sembrano fortemente dipendenti dai metodi utilizzati per la misurazione dell’affinità del ligando. Ad esempio, è stato riportato che i PBP, come quelli delle falene P. xylostella e Eogystia hippophaecolus, possono legare entrambi i componenti del feromone sessuale e gli analoghi (Sun et al., 2013a; Hu et al., 2018). Ciò suggerisce che i giocatori a valle come ORs potrebbero migliorare la specificità e la sensibilità della ricezione odorante., Prove recenti sostengono che la co-espressione di PBPS e PRs può aumentare la sensibilità verso i feromoni. Ad esempio, combinazioni multiple da PR1-4 e PBP1-4 sono state utilizzate per testare la loro risposta ai componenti del feromone sessuale della falena Chilo suppressalis (Chang et al., 2015). Gli autori hanno riscontrato un aumento significativo della sensibilità di risposta verso (Z)-11-esadecenale quando PR4 e PR6 sono stati co-espressi con PBP4., Anche se l’interazione di queste proteine potrebbe sorgere un nuovo livello di ricerca come obiettivi di controllo dei parassiti, il diverso abbinamento di PRs e PBP far luce sulla complessità del sistema olfattivo negli insetti, rendendo l’approccio un compito difficile per un ampio insieme di composti e proteine da testare. Nonostante quanto sopra, gli OBP degli insetti sono di piccole dimensioni molecolari con una facile produzione di proteine ricombinanti, il che li rende bersagli preferiti per studi strutturali e screening di legame rapido., Ad esempio, lo screening del ligando con OBPs potrebbe consentire l’identificazione di proprietà chimiche per un migliore legame, come la lunghezza della catena, il volume molecolare, i gruppi funzionali e l’insaturazione del legame. Questi, combinati con nuovi metodi di previsione della struttura proteica utilizzati nella progettazione di farmaci e anticorpi medici, come la modellazione dell’omologia, le simulazioni dinamiche e l’attracco molecolare, potrebbero posizionare gli OBP degli insetti in una posizione preferita rispetto agli OR come obiettivi per lo sviluppo di agenti di controllo nella gestione dei parassiti.
Gli OR degli insetti sembrano più specificamente sintonizzati sugli odoranti rispetto agli OBP., La maggiore specificità mostrata da ORs e la possibilità di attivazione/inibizione di recettori specifici per un determinato comportamento rendono queste proteine come bersagli attraenti per manipolare i comportamenti dei parassiti. La fattibilità dell’inibizione del complesso ORx/ORco o ORco da parte degli antagonisti comprende una strategia promettente per interrompere il comportamento specifico degli insetti, come l’accoppiamento tramite i recettori dei feromoni sessuali. Tuttavia, la mancanza di informazioni strutturali è il collo di bottiglia nell’utilizzo di OR di insetti come obiettivi per le previsioni di attività semiochimiche., Le tabelle 1, 2 riassumono il numero di OBPS e OR che sono stati identificati nelle specie di insetti mediante trascrittoma (cioè RNA-seq) e sequenziamento del genoma. La maggior parte degli insetti studiati finora hanno almeno due volte OR rispetto a OBPS secondo gli studi sul genoma. Inoltre, vi è una vasta espansione di OR negli insetti sociali dell’ordine degli imenotteri come l’ape mellifera A. con 170 OR (Weinstock et al., 2006), e le formiche Solenopsis invicta e Cerapachys biroi con 400 e 506 OR, rispettivamente (Wurm et al., 2011; Oxley et al., 2014)., Allo stesso modo, l’espansione OR è evidente anche in alcuni parassiti agricoli, come il coleottero della farina rossa T. castaneum con 265 OR rispetto a 47 OBPs (Richards et al., 2008). Questo rende un compito impegnativo per l’obiettivo O la selezione insieme alla difficoltà per l’espressione funzionale delle proteine transmembrana come ORs al fine di schermare un gran numero di ligandi. Un’approssimazione delle proprietà importanti sia in OBPs che in RUP è riassunta nella tabella 3.
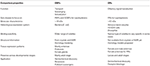
Tabella 3., Confronto approssimativo di insetti ORs e OBPS in base alle proprietà.
Ulteriori prospettive
La caratterizzazione funzionale degli OR degli insetti e il loro comprovato ruolo nell’olfatto degli insetti hanno fatto luce sulla sensibilità e la specificità di queste proteine specifiche per gli insetti. Questi progressi miglioreranno ulteriormente la loro fattibilità come obiettivi di controllo dei parassiti attraverso la comprensione dei meccanismi di riconoscimento molecolare e delle interazioni combinatorie con gli OBP., D’altra parte, l’attuale massiccio sforzo nell’identificazione e caratterizzazione vincolante di OBPS in diverse specie di insetti importanti agricoli continuerà e fornirà ulteriori informazioni sulle loro funzioni nella fisiologia degli insetti. Pertanto, questa revisione propone come vantaggio principale per OBPS rispetto alle OR, la disponibilità di strutture di cristallo 3D e NMR, che con approcci a valle, come la modellazione omologica (quando necessario), l’attracco molecolare e la dinamica molecolare, raffinerebbero la ricerca di sostanze chimiche bioattive., Quest’ultimo in complemento con la misurazione dell’affinità dei ligandi accelererà lo studio degli OBP degli insetti da riconsiderare come obiettivi per la scoperta semiochimica e gli strumenti per progettare i super-ligandi nella gestione del controllo dei parassiti.
L’aspetto e lo sviluppo della resistenza agli insetticidi nei parassiti degli insetti hanno portato alla ricerca intensiva sull’olfatto degli insetti e sui meccanismi coinvolti nell’elaborazione neurale. È ben noto che un certo numero di recettori ed enzimi nel SNC degli insetti sono gli obiettivi per lo sviluppo della resistenza agli insetticidi (Figura 1)., È stato dimostrato che l’acetilcolinesterasi (AChE) in forma solubile fornisce la resistenza agli insetticidi organo-fosforo e carbammato, agendo come bioscavengers (Lee et al., 2015). Allo stesso modo, molteplici meccanismi di resistenza agli insetticidi sono stati dimostrati nell’afide M. persicae, coinvolgendo carbossilesterasi, canali del sodio, acido γ-amminobutirrico (GABA) e nAChR (Bass et al., 2014)., Come componenti importanti nel sistema nervoso periferico degli insetti e attori chiave nei comportamenti degli insetti, sia gli OPP degli insetti che gli OR rappresentano obiettivi alternativi per l’identificazione di composti con attività semiochimica (o effetto agonista) e strumenti per progettare forti antagonisti per migliorare le risposte comportamentali desiderate dei parassiti degli insetti e ridurre l’uso di insetticidi e la successiva resistenza.
Autore Contributi
HV ha scritto sezioni sulla struttura di OBP e ORs, tabelle sviluppate e figura., J-JZ ha concepito l’idea per l’articolo di revisione, ha scritto la sezione principale, come l’introduzione, la funzione e la struttura OBPs e il confronto OR-OBP.
Dichiarazione sul conflitto di interessi
Gli autori dichiarano che la ricerca è stata condotta in assenza di relazioni commerciali o finanziarie che potrebbero essere interpretate come un potenziale conflitto di interessi.
Ringraziamenti
Gli autori ringraziano FONDECYT 3170433., J-JZ è grato per il sostegno finanziario da Northeast Normal University e Jilin University, Cina per il suo congedo sabbatico per studiare in Cina. Rothamsted Research riceve una borsa di studio con ulteriori finanziamenti da Biotechnology and Biological Sciences (BBSRC), Regno Unito.
Gonzalez, F., Witzgall, P., e Walker, WB (2017). I trascrittomi antennali di tre falene tortricide rivelano presunti recettori chemosensoriali conservati per segnali olfattivi sociali e di habitat. Sic. Rep. 7, 1-12. doi: 10.,1038/srep41829
PubMed Abstract | CrossRef Full Text/Google Scholar
Kaissling, K. E. (2013). La cinetica delle risposte olfattive potrebbe dipendere in gran parte dall’interazione odorante-recettore e dalla disattivazione odorante postulata per i rivelatori di flusso. J. Comp. Physiol. Neuroetolo. Comportamento neurale sensoriale. Physiol. 199, 879–196. doi: 10.1007 / s00359-013-0812-z
PubMed Abstract / CrossRef Full Text / Google Scholar