Progettazione e costruzione di randomizzati librerie per diluire Cas9 PAM preferenze
librerie PAM contenente randomizzati sequenze di DNA immediatamente a valle di una sequenza di DNA complementari per il distanziale di una guida di RNA sono stati generati e utilizzati per determinare empiricamente la PAM di riconoscimento di Tipo II Cas9 endonucleasi (Fig. 1)., Con la sequenza dell’obiettivo del distanziatore del RNA della guida che è fissa, le basi randomizzate servono da substrato per la lettura diretta della specificità di PAM dell’endonucleasi Cas9. Sequenze randomizzate sono state introdotte in un vettore di DNA plasmidico nella regione PAM di una sequenza bersaglio protospacer dimostrando perfetta omologia al distanziatore guida RNA T1 (CGCUAAAGAGGAAGAGGACA). Sono state generate due librerie che aumentano di dimensioni e complessità da cinque coppie di basi randomizzate (1.024 potenziali combinazioni PAM) a sette coppie di basi randomizzate (16.384 potenziali combinazioni PAM)., La randomizzazione della libreria 5 bp è stata introdotta attraverso la sintesi di un singolo oligonucleotide contenente cinque residui casuali. L’oligonucleotide a filamento singolo è stato convertito in un modello a doppio filamento mediante PCR (file aggiuntivo 1: Figura S1A), clonato nel vettore plasmidico (file aggiuntivo 1: Figura S1B) e trasformato in E. coli come descritto nella sezione Metodi., Per garantire la casualità ottimale nella libreria 7 bp PAM, la dimensione e la complessità della libreria sono state ridotte sintetizzando quattro oligonucleotidi contenenti ciascuno sei residui casuali più un settimo residuo fisso comprendente G, C, A o T, rispettivamente. Ciascuno dei quattro oligonucleotidi è stato convertito separatamente in DNA a doppio filamento, clonato nel vettore pTZ57R / T come descritto nella sezione Metodi e trasformato in E. coli come descritto per la libreria 5 bp., Dopo la trasformazione, il DNA plasmidico è stato recuperato e combinato da ciascuna delle quattro librerie PAM 6 bp per generare una libreria PAM 7 bp randomizzata comprendente 16.384 possibili combinazioni PAM. Per entrambe le librerie, l’incorporazione della casualità è stata convalidata mediante sequenziamento profondo; esaminando la composizione nucleotidica in ogni posizione della regione PAM utilizzando una matrice di frequenza di posizione (PFM ) (Sezione Metodi e) (File aggiuntivo 1: Figura S2A e B)., La distribuzione e la frequenza di ciascuna sequenza PAM nella libreria PAM randomizzata da 5 bp e 7 bp sono mostrate nel file aggiuntivo 1: Figure S3 e S4, rispettivamente.
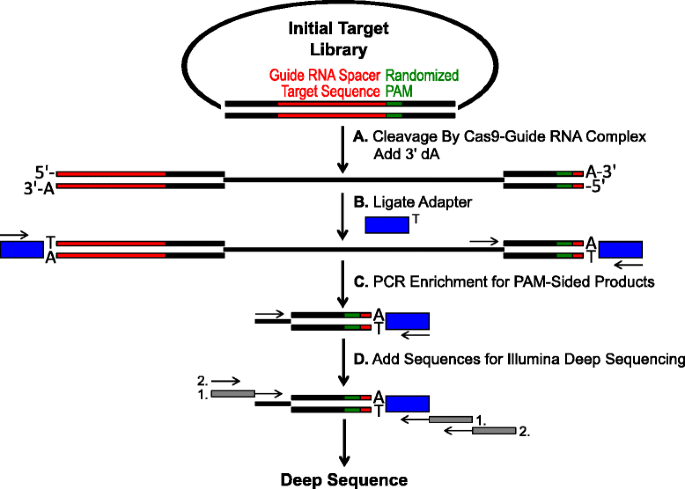
Schema per l’identificazione delle preferenze PAM mediante scissione Cas9 in vitro. una libreria plasmidica iniziale con PAM randomizzato (green box) viene scissa con il complesso Cas9 e vengono aggiunti sbalzi dA 3′. gli adattatori di b con 3 ‘ sporgenza di dT (scatola blu) sono legati ad entrambe l’estremità del prodotto di scissione., c Primer sono utilizzati per arricchire per PAM-sided cleaved prodotti da PCR., d Dopo la PCR arricchimento, i frammenti di DNA sono purificati e Illumina compatibile con le ancore e i codici a barre sono ‘coda-on’ attraverso due cicli di PCR (caselle di colore grigio) e Illumina in profondità sequenziato
Analizzando Cas9 PAM preferenze
Gli studi librerie PAM descritto nella sezione precedente sono stati sottoposti a digestione in vitro con diverse concentrazioni di ricombinante Cas9 proteina precaricato con guida RNA per dosaggio Cas9 endonucleasi PAM preferenze in modo dose-dipendente., Dopo la digestione con i complessi di ribonucleoproteina (RNP) dell’RNA della Cas9-guida, le combinazioni di sequenza di PAM dalla libreria randomizzata di PAM che hanno sostenuto la scissione sono state catturate legando gli adattatori alle estremità libere delle molecole del DNA del plasmide scisse dal complesso dell’RNA della Cas9-guida (Fig. 1 bis e b). Per promuovere la legatura efficiente e la cattura delle estremità scisse, il taglio del DNA a doppio filamento smussato generato dalle endonucleasi Cas9 è stato modificato per contenere uno sbalzo dA 3′ e gli adattatori sono stati modificati per contenere uno sbalzo DT complementare 3′., Per generare quantità sufficienti di DNA per il sequenziamento, i frammenti di DNA che ospitavano la sequenza PAM che supportava la scissione sono stati PCR amplificati utilizzando un primer nell’adattatore e un altro direttamente adiacente alla regione PAM (Fig. 1c). Le librerie Cas9 PAM amplificate per PCR risultanti sono state convertite in modelli di ampli-seq (Fig. 1d) e sequenziato in profondità a lettura singola dal lato adattatore dell’amplicone., Per garantire una copertura adeguata, le librerie PAM Cas9 sono state sequenziate ad una profondità almeno cinque volte maggiore della diversità nella libreria PAM randomizzata iniziale (5.120 e 81.920 letture per le librerie randomizzate PAM 5 e 7 bp, rispettivamente). Le sequenze PAM sono state identificate dai dati di sequenza risultanti selezionando solo quelle letture contenenti una perfetta corrispondenza di sequenza 12 nt che fiancheggia entrambi i lati della sequenza PAM 5 o 7 nt (a seconda della libreria PAM randomizzata utilizzata); catturando solo quelle sequenze PAM risultanti dal perfetto riconoscimento e scissione del sito bersaglio dell’RNA Cas9-guida., Per compensare il pregiudizio intrinseco nelle librerie PAM randomizzate iniziali, la frequenza di ogni sequenza PAM è stata normalizzata alla sua frequenza nella libreria iniziale. Poiché il test descritto qui cattura direttamente le sequenze PAM clivabili Cas9, è stata utilizzata la modellazione probabilistica per calcolare il consenso PAM per ciascuna proteina Cas9. Ciò è stato ottenuto valutando la probabilità di trovare ciascun nucleotide (G, C, A o T) in ogni posizione della sequenza PAM indipendentemente utilizzando una matrice di frequenza di posizione (PFM) (sezione Metodi e )., Le probabilità risultanti sono state quindi visualizzate come un WebLogo .
Per esaminare la propensione ai falsi positivi nel test, è stata omessa l’aggiunta di complessi Cas9 RNP nella fase di digestione (Fig. 1a) e il test è stato eseguito attraverso la fase di arricchimento della PCR (Fig. 1c). Come mostrato nel file aggiuntivo 1: Figura S5A, nessun prodotto di amplificazione è stato rilevato nell’assenza di complessi di RNA Cas9-guida. Pertanto, indicando che l’incidenza di falsi positivi è bassa e non contribuisce in modo significativo ai risultati del test.,
Preferenze PAM di Streptococcus pyogenes e Streptococcus thermophilus (sistemi CRISPR3 e CRISPR1) Proteine Cas9
Al fine di convalidare il test, sono state esaminate le preferenze PAM di Streptococcus pyogenes (Spy) e Streptococcus thermophilus CRISPR3 (Sth3) proteine Cas9, i cui requisiti di sequenza PAM sono stati precedentemente riportati. I digerimenti in vitro sono stati effettuati con 1 µg (5,6 nM) della libreria PAM randomizzata da 5 bp a due concentrazioni, 0,5 e 50 nM, di complessi Spy o Sth3 Cas9 preassemblati, CRRNA e tracrRNA RNP per 1 h in un volume di reazione di 100 µL., In base alla loro frequenza nella libreria PAM randomizzata a 5 bp, le sequenze PAM Spy e Sth3 Cas9 (NGG e NGGNG, rispettivamente) erano a concentrazioni finali di 0,40 nM e 0,11 nM nella digestione, rispettivamente. I membri della libreria PAM randomizzata che conteneva sequenze PAM che supportavano la scissione sono stati catturati e identificati come descritto nella sezione precedente. Come controllo negativo, la libreria PAM randomizzata non salvata iniziale era soggetta a sequenziamento e analisi PFM accanto a quelle librerie esposte a complessi RNP Cas9., Come mostrato nel file aggiuntivo 1: Figura S5B e C, non esistono preferenze di sequenza in assenza di digestione complessa Cas9 RNP come evidente da una distribuzione quasi perfetta di ciascun nucleotide in ogni posizione del PAM nella tabella PFM e dalla mancanza di contenuto informativo nel WebLogo per il controllo. Questo è in stark constrast con Fig. 2a e b che illustra la composizione delle sequenze derivate da librerie digerite con complessi Spy e Sth3 Cas9 RNP. Esame dei WebLogos derivati da PFM (Fig., 2a e b) rivelano anche la presenza delle preferenze PAM canoniche per le proteine Spy e Sth3 Cas9, NGG e NGGNG , rispettivamente. Sebbene le preferenze PAM riportate per le proteine Spy e Sth3 Cas9 siano osservate sia nei digestori da 0,5 nM che da 50 nM, vi è un generale ampliamento della specificità nelle condizioni di digest da 50 nM. Questo è più evidente in posizione 2 per la proteina Spy Cas9 dove la frequenza di un residuo non canonico aumenta drammaticamente (Fig. 2 bis)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
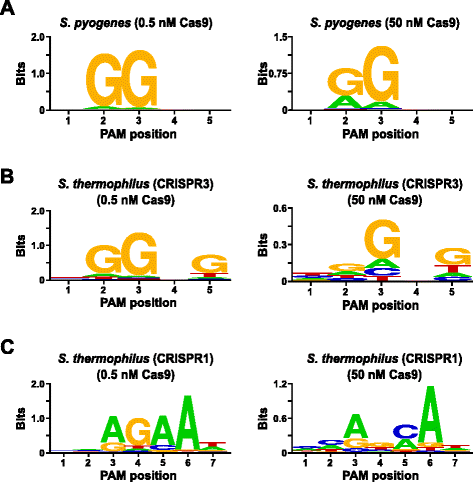
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., La frequenza dei nucleotidi in ciascuna posizione PAM è stata calcolata indipendentemente utilizzando una matrice di frequenza di posizione (PFM) e tracciata come WebLogo
Un’ulteriore validazione del test è stata condotta esaminando le preferenze PAM per la proteina Cas9 Streptococcus thermophilus CRISPR1 (Sth1) la cui specificità PAM è stata riportata estendersi a 7 bp . Utilizzando 1 µg (5,6 nM) della libreria PAM randomizzata da 7 bp come modello, le digestioni di RNA Sth1 Cas9-guide sono state eseguite a due concentrazioni, 0,5 nM e 50 nM, del complesso RNP come descritto sopra., Come controlli, i complessi Spy e Sth3 Cas9 RNP sono stati utilizzati anche per digerire la libreria PAM randomizzata 7 bp ma solo a una singola concentrazione di complesso RNP di 0,5 nM. Sulla base della frequenza nella libreria PAM randomizzata 7 bp, le sequenze PAM precedentemente riportate per Sth1 (NNAGAAW), Spy (NGG) e Sth3 (NGGNG) erano rispettivamente a concentrazioni finali di 0,01 nM, 0,22 nM e 0,05 nM., Come mostrato nel file aggiuntivo 1: Figura S6A e B, le preferenze PAM per le proteine Spy e Sth3 Cas9 generate utilizzando la libreria 7 bp erano quasi identiche a quelle prodotte con la libreria 5 bp fornendo una forte evidenza per la riproducibilità del test. Anche le preferenze PAM per la proteina Cas9 Sth1 corrispondevano strettamente a quelle precedentemente riportate, NNAGAAW, alla concentrazione del complesso di RNA Cas9-guida di 0,5 nM (Fig. 2 quater)., Simile alle proteine Spy e Sth3 Cas9, Sth1 Cas9 era in grado di scindere un insieme più diversificato di sequenze PAM nelle reazioni contenenti una maggiore concentrazione di complesso RNA Cas9-guida (50 nM), la più sorprendente era la marcata perdita del requisito del residuo G alla posizione 4 e la preferenza quasi uguale per un C e un bp alla posizione 5 (Fig. 2 quater). Ciò ha comportato un consenso PAM diverso da quello ottenuto a concentrazioni più basse.,
Per esaminare se la specificità PAM è indipendente dal tipo di RNA guida, crRNA duplex:tracrRNA o sgRNA , Spy, Sth3 e Sth1 Le preferenze PAM Cas9 sono state esaminate anche utilizzando un complesso binario Cas9 e sgRNA RNP. La digestione è stata effettuata ad una singola concentrazione complessa di RNP di 0,5 nM e l’analisi delle preferenze PAM è stata eseguita come descritto sopra. Come mostrato nel file aggiuntivo 1: Figura S7A, B e C, le preferenze PAM erano quasi identiche indipendentemente dal tipo di RNA guida utilizzato; o un crRNA:tracrRNA duplex o sgRNA., Inoltre, per confermare che la specificità PAM non è fortemente influenzata dalla composizione del DNA bersaglio o della sequenza di distanziatori, la sequenza sul lato opposto della libreria randomizzata 5 o 7 bp è stata mirata alla scissione con un distanziatore diverso; T2-5 (UCUAGAUAGAUUACGAAUUC) per la libreria 5 bp o T2-7 (CCGGCGACGUUGGGUCAACU) per la libreria 7 bp. Le proteine Spy e Sth3 Cas9 precaricate con SGRNA mirate alla sequenza T2 sono state utilizzate per interrogare la libreria PAM randomizzata da 5 bp mentre i complessi Sth1 Cas9-T2 sgRNA sono stati utilizzati per digerire la libreria PAM randomizzata da 7 bp., Le preferenze PAM sono state analizzate come descritto sopra. Le preferenze PAM per tutte e 3 le proteine Cas9 erano quasi identiche indipendentemente dal distanziatore e dalla sequenza del DNA bersaglio (file aggiuntivo 1: Figura S8A, B e C).
Identificazione delle preferenze di sgRNA e PAM per la proteina Cas9 di Brevibacillus laterosporus
Per esaminare empiricamente le preferenze di PAM per una proteina Cas9 la cui PAM non era definita, è stato identificato un locus CRISPR-Cas di tipo II-C non caratterizzato dal ceppo Brevibacillus laterosporus SSP360D4 (Blat) ricercando database interni DuPont Pioneer per, Il locus (circa 4,5 kb) conteneva un gene cas9 in grado di codificare un polipeptide 1,092, un array CRISPR comprendente sette unità di distanziatore di ripetizione appena a valle del gene cas9 e una regione di codifica tracrRNA situata a monte del gene cas9 con omologia parziale alle ripetizioni dell’array CRISPR (Fig. 3 bis). La lunghezza della ripetizione e del distanziatore (36 e 30 bp, di conseguenza) è simile ad altri sistemi CRISPR-Cas di tipo II con cinque delle otto ripetizioni contenenti 1 o 2 mutazioni bp (Fig. 3b e file aggiuntivo 1: Figura S9)., Altri geni che si trovano tipicamente in un locus CRISPR-Cas di tipo II erano troncati (cas1) o mancanti (Fig. 3 bis).
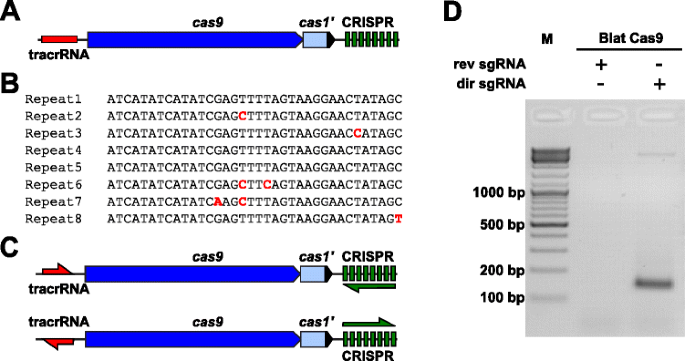
Identificazione di elementi CRISPR-Cas di tipo II nel sistema CRISPR-Cas di Brevibacillus laterosporus SSP360D4. a Un’illustrazione della regione genomica del DNA dal sistema CRISPR-Cas di tipo II da Brevibacillus laterosporus SSP360D4. b Confronto delle sequenze di ripetizione dell’array CRISPR di tipo II identificate in Brevibacillus laterosporus SSP360D4., c Gli scenari trascrizionali degli array di tracrRNA e CRISPR “diretti” e “inversi” per il sistema CRISPR-Cas di tipo II da Brevibacillus laterosporus SSP360D4. d Un gel di agarosio con prodotti di reazione, che indica che solo la ‘diretta’ sgRNA (dir sgRNA), ma non il ‘reverse’ sgRNA (rev sgRNA) supporto plasmide biblioteca scissione in combinazione con il Cas9 endonucleasi provenienti da Brevibacillus laterosporus SSP360D4
La guida di RNA requisito per il Blat Cas9 di proteine è stata determinata mediante la generazione di due sgRNA varianti., Queste varianti sono state generate per tenere conto di entrambi i possibili scenari di espressione sense o anti-sense dell’array tracrRNA e CRISPR (Fig. 3c) e utilizzato per sondare quale scenario di espressione supportava l’attività di scissione di Blat Cas9 nella libreria PAM randomizzata. Gli RNA a guida singola sono stati progettati identificando prima i confini delle molecole di tracrRNA putative analizzando le regioni che erano parzialmente complementari al 22 nt 5 ‘ terminale della ripetizione (anti-ripetizione)., Successivamente, per determinare l’estremità 3 ‘ del tracrRNA, sono state utilizzate possibili strutture secondarie e terminatori per prevedere la regione di terminazione nel frammento a valle. Ciò è stato ottenuto mediante screening per la presenza di sequenze di terminazione simili a Rho indipendenti nel DNA che circonda l’anti-ripetizione simile a quella descritta in Karvelis et al. , convertendo il DNA circostante in sequenza di RNA ed esaminando le strutture risultanti usando UNAfold ., Gli SGRNA risultanti sono stati progettati per contenere un segnale di riconoscimento di iniziazione della trascrizione della polimerasi T7 all’estremità 5′ seguito da una sequenza di riconoscimento target 20 nt, 16 nt di ripetizione crRNA, 4 nt self-folding hairpin loop e sequenza anti-ripetizione complementare alla regione di ripetizione del crRNA seguita dalla restante parte 3’ del tracrRNA putativo. La variante sgRNA che contiene un tracrRNA putativo trascritto nella stessa direzione del gene cas9 (Fig. 3c) è definito sgRNA ‘diretto‘, mentre lo sgRNA contenente il tracrRNA trascritto nella direzione opposta uno sgRNA’ inverso’., Cinquanta nM di Blat Cas9 sgRNA RNP complex, pre-caricati rispettivamente con SGRNAS “diretto” o “inverso”, sono stati incubati con 1 µg (5,6 nM) della libreria PAM randomizzata 7 bp. Dopo la digestione della libreria e l’aggiunta di sporgenze dA 3′, gli adattatori sono stati legati e i prodotti di scissione sono stati PCR amplificati (Fig. 1). L’analisi dei prodotti di reazione mediante elettroforesi su gel di agarosio ha rivelato che lo sgRNA “diretto”, ma non lo sgRNA “inverso”, supportava la scissione della libreria plasmidica (Fig. 3d). La sequenza e la struttura secondaria prevista dello sgRNA “diretto” sono mostrate nel file aggiuntivo 1: Figura S10.,
Dopo aver determinato l’RNA guida appropriato per Blat Cas9, l’identificazione PAM è stata eseguita in modo simile a quella descritta sopra per le proteine Spy, Sth3 e Sth1 Cas9 contro la libreria PAM randomizzata da 7 bp con due concentrazioni, 0,5 e 50 nM, del complesso sgRNA RNP Blat Cas9 “diretto” preassemblato. Come mostrato in Fig. 4a, il consenso PFM WebLogo PAM per la proteina Blat Cas9 nelle condizioni di digest di 0,5 nM era NNNNCND (N = G, C, A o T; D = A, G o T) con una forte preferenza per una C alla posizione 5 della sequenza PAM., Una moderata preferenza per una A è stata osservata alla posizione 7 e lievi preferenze per una C o T alla posizione 4 e G, C o A su T alla posizione 6 sono state notate anche esaminando attentamente la tabella PFM (File aggiuntivo 1: Figura S11). Analogamente alle proteine Spy, Sth3 e Sth1 Cas9, la specificità PAM si allarga all’aumentare della concentrazione del complesso Cas9-sgRNA. Ciò è più evidente in posizione 5 dove una percentuale maggiore di sequenze PAM contenenti una scissione di supporto residuo A 50 nM rispetto alle condizioni di digest 0.5 nM.
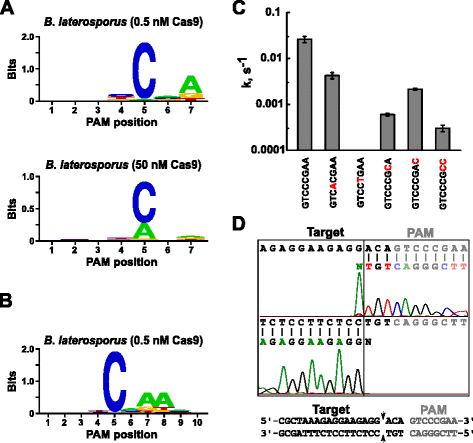
Preferenze PAM e posizioni di scissione dell’enzima Brevibacillus laterosporus SSP360D4 (Blat) Cas9. Preferenze Blat Cas9 PAM quando 1 µg di DNA libreria è stato scisso con 0,5 nM o 50 nM complesso Cas9-sgRNA (a), esteso alla posizione 10 spostando il bersaglio protospacer di 3 bp (b). La frequenza dei nucleotidi in ogni posizione PAM è stata calcolata indipendentemente utilizzando una matrice di frequenza di posizione (PFM) e tracciata come WebLogo ., c Tassi di scissione di substrati di DNA plasmidico supercoiled contenenti mutazioni (mostrato in rosso) nella sequenza GTCCCGAA PAM. Tutti i punti dati sono valori medi di ≥3 esperimenti indipendenti. Le barre di errore sono fornite come sequenziamento del Run-off S. D. d da entrambe le direzioni senso e anti-senso del DNA plasmidico scisso con Blat Cas9
Poiché Blat Cas9 può accettare qualsiasi base nelle prime tre posizioni della sua sequenza PAM (Fig. 4a), il distanziatore T1 è stato spostato da tre nucleotidi nella direzione 5′ per consentire l’identificazione PAM da estendere da 7 a 10 bp., Il distanziatore T1 spostato, T1-3 (AAACGCUAAAGAGGAAGAGG), è stato incorporato nello sgRNA “diretto” Blat e l’identificazione PAM è stata eseguita come descritto in precedenza per le proteine Spy, Sth3, Sth1 e Blat Cas9. L’analisi delle preferenze PAM ha rivelato che la specificità PAM per Blat Cas9 può essere estesa alla posizione 8 dove c’è una preferenza moderata per un ulteriore A (Fig. 4 ter).
La specificità del PAM per Blat Cas9 è stata confermata generando plasmidi per contenere mutazioni nei residui più conservati del PAM (Fig. 4 quater)., La sostituzione del nucleotide C alla posizione 5 ha abolito la scissione del DNA plasmidico confermando il suo ruolo chiave nel riconoscimento Blat Cas9 PAM. La sostituzione di un nucleotide nelle posizioni 7 e 8 ha ridotto significativamente (43× e 12×, rispettivamente) il tasso di scissione del plasmide supercoiled indicando anche l’importanza di questi nucleotidi nel riconoscimento Blat Cas9 PAM.,
Per identificare le posizioni di scissione del DNA bersaglio per la proteina Blat Cas9, un plasmide contenente una regione di 20 bp corrispondente al distanziatore T1 seguito da una sequenza PAM, GTCCCGAA, che rientra nel consenso PAM per Blat Cas9, NNNNCNDD, è stato generato e digerito con Blat Cas9-guida RNA ribonucleoproteina complesso. Il sequenziamento diretto del DNA è stato utilizzato per determinare le estremità della molecola di DNA lineare generata dal complesso Blat Cas9 RNP. I risultati della sequenza hanno confermato che la scissione del DNA plasmidico si è verificata nel protospacer 3 nt 5 ‘ della sequenza PAM (Fig., 4d) simile a quello osservato per le proteine Spy, Sth3 e Sth1 Cas9 .
Nell’editing del genoma di planta utilizzando Blat Cas9 e sgRNA
A seguito della delucidazione delle preferenze sgRNA e PAM per Blat Cas9, sono state generate cassette di espressione Cas9 e sgRNA ottimizzate per il mais per i test in planta come precedentemente descritto per il gene S. pyogenes cas9 e sgRNA . In breve, il gene Blat cas9 è stato ottimizzato per il codone di mais e l’introne 2 del gene ST-LSI della patata è stato inserito per interrompere l’espressione in E. coli e facilitare lo splicing ottimale in planta (File aggiuntivo 1: Figura S12)., Localizzazione nucleare della proteina Blat Cas9 nelle cellule di mais è stato facilitato dall’aggiunta di entrambi amminico e carbossile-terminale segnali posizioni nucleari, SV40 (MAPKKKRKV) e Agrobacterium tumefaciens VirD2 (KRPRDRHDGELGGRKRAR), rispettivamente (File aggiuntivo 1: Figura S12). Il gene Blat cas9 è stato espresso in modo costitutivo nelle cellule vegetali collegando il cas9 ottimizzato a un promotore di ubiquitina di mais e al terminatore pinII in un vettore di DNA plasmidico., Per conferire un’espressione efficiente dello sgRNA nelle cellule di mais, un promotore e un terminatore della polimerasi III U6 di mais (TTTTTTTT) sono stati isolati e fusi rispettivamente alle estremità 5′ e 3′ di una sequenza di DNA modificata che codifica Blat sgRNA (File aggiuntivo 1: Figura S13). Lo sgRNA Blat modificato conteneva due modifiche rispetto a quello utilizzato negli studi in vitro; un’alterazione da T a G alla posizione 99 e una modifica da T a C alla posizione 157 dello sgRNA (File aggiuntivo 1: Figura S13). Le modifiche sono state introdotte per rimuovere i potenziali segnali prematuri di terminazione della polimerasi III U6 nel Blat sgRNA., Alterazioni introdotte per avere un impatto minimo sulla struttura secondaria dello sgRNA rispetto alla versione utilizzata negli studi in vitro (dati non mostrati).
Per confrontare con precisione l’efficienza mutazionale risultante dalla riparazione imperfetta non omologa di end-joining (NHEJ) delle rotture a doppio filamento di DNA (DSB) derivanti dalla scissione di Spy e Blat Cas9, i siti di destinazione genomici identici di protospacer sono stati selezionati identificando obiettivi con PAM compatibili con Spy e Blat Cas9, NGGNCNDD., Sequenze di distanziatori identiche sono state selezionate per Blat e Spy Cas9 catturando la sequenza da 18 a 21 nt immediatamente a monte del PAM. Per garantire un’espressione ottimale della polimerasi III U6 e non introdurre una mancata corrispondenza all’interno del distanziatore sgRNA, tutte le sequenze target sono state selezionate per terminare naturalmente in un G alla loro estremità 5′. Gli obiettivi sono stati identificati e selezionati nell’esone 1 e 4 del gene della fertilità del mais Ms45 e in una regione a monte del gene liguleless-1 del mais.,
L’attività mutazionale di Blat Cas9 nel mais è stata esaminata trasformando biolisticamente embrioni immaturi di mais di 10 giorni con vettori di DNA contenenti geni cas9 e sgRNA. Blat e l ” equivalente Spy Cas9 e sgRNA vettori di espressione sono stati introdotti in modo indipendente in mais Hi-Type II IMEs dalla trasformazione pistola particella simile a quella descritta in . Poiché la trasformazione della pistola di particelle può essere altamente variabile, una cassetta di espressione del DNA del marcatore visivo, Ds-Red, è stata anche co-consegnata con i vettori di espressione Cas9 e sgRNA per aiutare nella selezione di IME uniformemente trasformati., In totale, tre repliche di trasformazione sono state eseguite su 60-90 IME e 20-30 delle IME più uniformemente trasformate da ciascuna replica sono state raccolte 3 giorni dopo la trasformazione. Il DNA genomico totale è stato estratto e la regione che circonda il sito bersaglio è stata amplificata da PCR e ampliconi sequenziati a una profondità di lettura superiore a 300.000. Le letture risultanti sono state esaminate per la presenza di mutazioni nel sito previsto di scissione rispetto agli esperimenti di controllo in cui la cassetta di espressione del DNA sgRNA è stata omessa dalla trasformazione. Come mostrato in Fig., 5a, mutazioni sono state osservate nel sito di scissione previsto per Blat Cas9 con i tipi più prevalenti di mutazioni che sono inserzioni o eliminazioni di coppie di basi singole. Modelli di riparazione simili sono stati osservati anche per la proteina Spy Cas9 (file aggiuntivo 1: Figura S14 e ). L’attività mutazionale per Blat Cas9 è stata robusta in due dei tre siti testati e ha superato quella della Spia Cas9 nel sito bersaglio Ms45 exon 4 di circa il 30% (Fig. 5 ter).
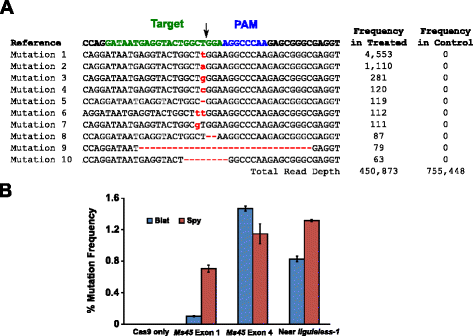
Brevibacillus laterosporus Cas9 promuove mutazioni NHEJ nel mais. a Top 10 tipi più prevalenti di mutazioni NHEJ rilevate con Blat Cas9 nell’esone 4 del gene Ms45. Una freccia nera indica il sito previsto di scissione; le mutazioni sono evidenziate in rosso; il carattere minuscolo indica un inserimento; ‘ – ’ indica una cancellazione. b Confronto delle frequenze di mutazione Spy e Blat Cas9 NHEJ in tre siti bersaglio identici protospacer nel mais. Le mutazioni di NHEJ sono state rilevate mediante sequenziamento profondo 3 giorni dopo la trasformazione., Le barre di errore rappresentano SEM, n = 3 trasformazioni di pistole di particelle. Solo Cas9 è il controllo negativo e rappresenta la frequenza media (in tutti e tre i siti target) di base delle mutazioni risultanti dall’amplificazione e dal sequenziamento della PCR