Selv om flyktige forbindelser med agonist aktivitet har vært skjermet mot Or, et bestemt kjemisk stoff med sterk agonist effekt på mygg, 2-(4-Ethyl-5-(pyridin-3-yl)-4-1,2,4-triazol-3-ylthio)-N-(4-ethylphenyl)acetamide (VUAA1), har åpnet innen forskning (Jones et al., 2011; Taylor et al., 2012). Senere studier av Taylor et al., (2012) bevis for VUAA1-avledet kjemikalier, for eksempel VUAA4, i stand til å øke sin agonist effekt ved 10-brett på ORco fra A. gambiae, H. virescens, og Harpegnathos saltator. Det er interessant at forfatterne rapporterte at enhver endring på amide substituenter vil føre til en fullstendig tap av agonist aktivitet. Dette gir nyttig innsikt i de strukturelle kravene i agonister og struktur-aktivitet forholdet mellom VUAA analogs og Or. Til slutt, til tross for forbedret agonist aktivitet av VUAA kjemikalier, sin relativt høy molekylvekt (367.47 g mol−1 for VUAA1) vs., flyktige agonister, som bombykol (238.42 g mol−1), gjør en direkte flyktige levering av VUAA noe som ikke er gjennomførbart. Med det i tankene, den søker for mindre strukturelle analoger representerer et interessant fokus på forskning.
Antagonisme På ORco Subunit
Sammen med studiet av VUAA-relaterte analogs, som kan fungere som sterke agonister, blokkering av ORco av antagonister har også dukket opp for å veilede semiochemicals og plantevernmiddel design., Dermed, en strukturell analog av VUAA1, VU0183254 (2-(4-Ethyl-5-furan-2-yl-4H-triazol-3-ylsulfanyl)-1-phenothiazin-10-yl-ethanone), som ble rapportert å hemme ORco svar, fungerer som allosteric modulator i A. gambiae og forstyrre anerkjennelse av agonister som eugenol av komplekse OR65/ORco (Jones et al., 2012). Andre VUAA-strukturelle analoger har også vært rapportert som antagonister. Et eksempel er N-,2-substituerte triazolothioacetamide forbindelser OLC3 og OLC12 som forstyrrer ORco svar på en liknende måte som i C. quinquefasciatus, A. gambiae, D. melanogaster, og O., nubilalis, noe som tyder på en bevart bindende nettstedet i ORco (Chen og Luetje, 2012). Vurderer hemming av ORco som en lovende strategi for å forstyrre atferd av insekter, ser det ut til at etterfølgende tiltak bør ta sikte på å forbindelser med lavere molekylvekt enn VUAA mekano-antagonister. For eksempel, OX1a (232 g mol−1), tryptamine (160.22 g mol−1) og isopropyl cinnamate (190.24 g mol−1) ble rapportert å ha antagonist effekt på ORco (Chen og Luetje, 2013, 2014; Tsitoura et al., 2015) med omtrent halvparten eller mindre molekylvekt enn VUAA1., Likevel, fremtidige bruken av disse antagonister bør studeres nøye, siden blokkering av bevart ORco kan påvirke ikke bare skadelige insekter, men også gunstig seg.
Foruten antagonist effekt analysert in vitro, bevis på atferdsmessige nivå støtter ideen om at strukturelle analoger av feromoner kan fungere som antagonister. For eksempel, Sellanes et al., (2010) rapporterte hemming av seksuell respons i honeydew møll Cryptoblabes gnidiella når den strukturelle analoger, (Z)-9-tetradecenyl formiat og (Z)-11-hexadecenyl formiat, ble lagt til syntetiske sex feromoner, (Z)-11-hexadecenal og (Z)-13-octadecenal, i vindtunnel-tester. Denne pheromone antagonist effekten ble senere bekreftet i feltet analyser, der fangst av C. gnidiella menn gikk ned i en dose-avhengig mønster. Pheromone antagonisme har også blitt rapportert for B. mandarina, en stamfar av B. mori (Daimon et al., 2012)., Deres funn underbygge bombykol som sex feromoner, og bombykal og bombykyl acetat som antagonister, som er sterkt hemmet tiltrekningen av menn i feltet til sex feromoner bombykol. Mer nylig, bevis av pheromone motsetningen ble rapportert for snuten møll Herpetogramma submarginale. Når (Z)-13-hexadecenol ble lagt til sin sex feromoner, (Z)-13-hexadecenyl acetat, signifikant redusert antall menn fanget i feltet (Yan et al., 2015)., Pheromone motsetningen synes basert på forskjeller i kjemisk funksjonell gruppe som alkoholer, aldehyder og estere avhengig av insektarter. Likevel, antagonist effekten av disse strukturelle analoger kan ikke være på grunn av ORco-hemming, men spesifisiteten av ORx å antagonister. En fersk studie antyder at OR16 av Helicoverpa armigera er i stand til å spesifikt gjenkjenner pheromone antagonist, (Z)-11-hexadecenol (Chang H. et al., 2017). Forfatterne støttet den spesifikke rollen OR16 vurderer at H., armigera kvinner slipper ut antagonist compound sammen med sin sex feromoner ((Z)-11-hexadecenal og (Z)-9-hexadecenal) som en strategi for å unngå ikke-optimal parring med umodne hanner. Utrolig, når OR16 ble slått ned av genom redigering teknikk CRISPR/Cas9 og H. armigera menn ble testet av elektrofysiologi og atferdsmessige analyser, ingen EAG respons ble registrert og menn prøvde å parre seg med umodne hunner.
Odorant Reseptorer vs., Bindende Proteiner: Fordeler og Ulemper for Insekt Pest Management
For tilfelle av OBPs, ligand spesifisitet og mekanismer for OBPs representerer kontroversielle aspekter, som virker sterkt avhengig av metoder som brukes for måling av ligand affinitet. For eksempel, det har blitt rapportert at Pbp, slik som de som er fra møll’ S. xylostella og Eogystia hippophaecolus, kan binde begge kjønn pheromone komponenter og analogs (Sun et al., 2013a; Hu et al., 2018). Dette tyder på at nedstrøms aktører som Or kunne øke spesifisiteten og sensitiviteten av odorant resepsjon., Nyere bevis støtter at co-uttrykk av Pbp-og PRs kan øke følsomheten mot feromoner. For eksempel, flere kombinasjoner fra PR1-4 og PBP1-4 ble brukt til å teste ut deres respons til sex feromoner komponenter av møll Chilo suppressalis (Chang et al., 2015). Forfatterne fant en signifikant økning i følsomheten for reaksjon mot (Z)-11-hexadecenal når PR4 og PR6 var co-uttrykt med PBP4., Selv om samspillet av disse proteinene kan oppstå et nytt nivå av forskning som skadedyrbekjempelse mål, ulike sammenkobling av PRs og Pbp kaste lys på kompleksiteten av olfactory system i insekter, noe som gjør tilnærming en vanskelig oppgave for et stort sett av forbindelser og proteiner for å teste. Til tross for det ovennevnte, insekt OBPs er av små-molekylære størrelse med enkel produksjon av rekombinante proteiner, noe som gjør dem favoritt mål for strukturelle studier og rask bindende screening., For eksempel, ligand screening med OBPs kan føre til identifisering av kjemiske egenskaper for bedre binding, som kjede lengde, molekylær volum, funksjonelle grupper, og bond unsaturation. Disse, kombinert med nye protein struktur prediksjon metoder som brukes i utformingen av medisinske stoffer og antistoffer, for eksempel homologi modellering, dynamikk simuleringer og molekylær-dokk, kunne plassere insekt OBPs i en favoritt posisjon over Or som mål for utvikling av kontroll agenter i pest management.
Insekt Or virke mer spesielt tilpasset for å odorants enn OBPs., Den høyere spesifisitet vist av Or og sjansen for aktivering/hemming av spesifikke reseptorer for en gitt adferd gjør disse proteiner som attraktive mål å manipulere pest atferd. Muligheten for hemming av enten ORx/ORco komplekse eller ORco av antagonister består av en lovende strategi for å forstyrre insekt bestemt atferd, for eksempel parring via kjønn pheromone reseptorer. Imidlertid, i mangel av strukturell informasjon som er flaskehalsen i bruk av insekt Or som mål for semiochemical aktivitet spådommer., Tabellene 1, 2 oppsummerer antallet OBPs og Or som har blitt identifisert i insektarter av transcriptome (dvs., RNA-seq) og genom-sekvensering. De fleste av insekter studert så langt har minst to ganger Or enn OBPs i henhold til genom studier. Videre er det en omfattende utvidelse av Or i sosiale insekter fra Hymenopteran for eksempel honey bee A. mellifera med 170 Or (Weinstock et al., 2006), og maurene Solenopsis invicta og Cerapachys biroi med 400 og 506 Or, henholdsvis (Wurm et al., 2011; Oxley et al., 2014)., Tilsvarende, ELLER ekspansjon er også tydelig i noen landbruket skadedyr, som for eksempel rødt mel bille T. castaneum med 265 Or i forhold til 47 OBPs (Richards et al., 2008). Dette gjør en krevende oppgave for mål ELLER utvalg sammen med vanskeligheter for den funksjonelle uttrykk for en av verdens proteiner slik som Or for å skjermen et stort antall av ligander. En tilnærming av viktige egenskaper i både OBPs og Or er oppsummert i Tabell 3.
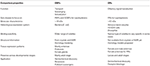
Tabell 3., Omtrentlig sammenligning av insekt Or og OBPs i henhold til egenskaper.
Videre Perspektiver
funksjonell karakterisering av insekt Or samt deres påvist roller i insekt luktesans har kaste lys på sensitivitet og spesifisitet av disse insekt-spesifikke proteiner. Disse fremskritt vil ytterligere forsterke deres gjennomførbarhet som skadedyrbekjempelse mål av forståelsen av molekylære anerkjennelse mekanismer og kombinasjoner interaksjoner med OBPs., På den annen side, gjeldende massiv innsats i identifisering og bindende karakterisering av OBPs i flere landbruket viktig insektarter vil fortsette og gi mer informasjon om hvilke funksjoner i insect physiology. Dermed, denne gjennomgangen foreslår som viktigste fordelen for OBPs over Or, tilgjengeligheten av 3D-crystal og NMR strukturer, som med nedstrøms tilnærminger, for eksempel homologi modellering (når nødvendig), molekylær-dokk og molekylær dynamikk, ville avgrense søk av bioaktive kjemikalier., Dette siste i komplettere med ligand affinitet måling vil akselerere studie av insekt OBPs å bli revurdert som mål for semiochemical discovery og verktøy for å designe super-ligander i skadedyrbekjempelse ledelse.
utseende og utvikling av insektmiddel motstand i insekt skadedyr har ført til en intens forskning på insekt luktesans og mekanismer som er involvert for nevrale behandling. Det er godt etablert at en rekke reseptorer og enzymer i insekt CNS er målene for insektmiddel utvikling av resistens (Figur 1)., Det har blitt demonstrert at acetylkolinesterase (AChE) i løselig form gir motstand til organo-fosfor og carbamate insektmidler, som opptrer som bioscavengers (Lee et al., 2015). På samme måte flere insektmiddel motstand mekanismer har vært påvist i aphid M. persicae, som involverer carboxylesterases, natrium-tv, γ-aminobutyric acid (GABA) og nAChR (Bass et al., 2014)., Som viktige komponenter i insekt periferi nerve systemet og sentrale aktører i insekt atferd, både insekt OBPs og Or representerer alternative mål for identifisering av forbindelser med semiochemical aktivitet (eller agonist effekt) og verktøy for å designe sterke motstandere for å forbedre ønsket atferdsmessige reaksjoner av insekt skadedyr og redusere bruken av plantevernmidler og påfølgende motstand.
Forfatter Bidrag
HV skrev avsnitt om OBP er struktur og Or, utviklet tabeller og figur., J-JZ unnfanget ideen til vurdering artikkelen, skrev den viktigste delen, som innledning, OBPs funksjon og struktur, samt ELLER-OBP sammenligningen.
interessekonflikt Uttalelse
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller finansielle forhold som kan oppfattes som en potensiell interessekonflikt.
Erkjennelsene
forfatterne ønsker å takke FONDECYT 3170433., J-JZ er takknemlig for den økonomiske støtten fra Nordøst Normal University og Jilin University, Kina for sine sabbatsår permisjon for å studere i Kina. Rothamsted Research mottar et stipend med ekstra finansiering fra Bioteknologi og Biologiske Fag (BBSRC), UK.
Gonzalez, F., Witzgall, P., og Walker, W. B. (2017). Antennal transcriptomes av tre tortricid møll avsløre mulige bevart chemosensory reseptorer for sosiale og habitat olfactory stikkord. Sci. Rep. 7, 1-12. doi: 10.,1038/srep41829
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar
Kaissling, K. E. (2013). Kinetikk for olfactory reaksjoner kan i stor grad avhenge av odorant-reseptor samhandling og odorant deaktivering postulerte for flux-detektorer. J. Red. Physiol. Neuroethol. Sensorisk Nerve Behav. Physiol. 199, 879-196. doi: 10.1007/s00359-013-0812-z
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar