Design and construction of randomized libraries for assaying Cas9 PAM preferences
PAM libraries containing randomized DNA sequenties immediately downstream of a DNA sequence complementary to the spacer of a guide RNA were generated and used to empirically determining the PAM recognition of type II Cas9 endonucleases (Fig. 1)., Met de opeenvolging van het de spacerdoel van gidsrna vast wordt gesteld, dienen de gerandomiseerde basissen als substraat voor de directe read-out van Cas9 endonuclease PAM specificiteit. De gerandomiseerde opeenvolgingen werden geà ntroduceerd in een plasmide DNA vector in het PAM gebied van een protospacer doelvolgorde die perfecte homologie aan de gidsrna spacer T1 (CGCUAAAGAGGAAGAGGACA) demonstreert. Twee bibliotheken die in omvang en complexiteit toenamen van vijf gerandomiseerde basisparen (1.024 potentiële PAM-combinaties) tot zeven gerandomiseerde basisparen (16.384 potentiële PAM-combinaties) werden gegenereerd., Randomisatie van de bibliotheek van 5 bp werd geà ntroduceerd door de synthese van één enkele oligonucleotide die vijf willekeurige residuen bevat. Het enkelstrengs oligonucleotide werd omgezet in een dubbelstrengs malplaatje door PCR( aanvullend dossier 1: Figuur S1A), gekloond in de plasmidevector (aanvullend dossier 1: Figuur S1B) en omgezet in E. coli zoals beschreven in de sectie methoden., Om optimale willekeur in de 7 BP PAM bibliotheek te verzekeren, werden de grootte en de complexiteit van de bibliotheek verminderd door vier oligonucleotiden samen te stellen die elk zes willekeurige residuen plus een zevende vaste residu bevatten die G, C, A, of T, respectievelijk bevatten. Elk van de vier oligonucleotiden werd afzonderlijk omgezet in dubbelstrengs DNA, gekloond in vector pTZ57R/T zoals beschreven in de sectie methoden en omgezet in E. coli zoals beschreven voor de bibliotheek 5 bp., Na transformatie werd plasmide-DNA teruggevonden en gecombineerd uit elk van de vier 6 bp PAM-bibliotheken om een gerandomiseerde 7 BP PAM-bibliotheek te genereren bestaande uit 16.384 mogelijke Pam-combinaties. Voor beide bibliotheken werd de opname van willekeur gevalideerd door deep sequencing; het onderzoeken van de nucleotidesamenstelling op elke positie van het PAM-gebied met behulp van een positiefrequentiematrix (PFM) (sectie methoden en ) (aanvullend dossier 1: Figuur S2A en B)., De distributie en frequentie van elke PAM sequentie in de 5 bp en 7 BP gerandomiseerde PAM bibliotheek worden weergegeven in aanvullend bestand 1: figuren S3 en S4, respectievelijk.
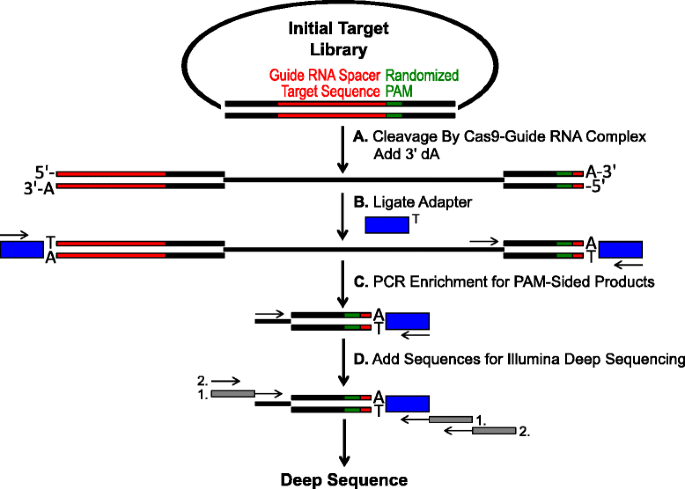
schema voor identificatie van PAM voorkeuren door Cas9 splitsing in vitro. een initiële plasmidebibliotheek met gerandomiseerde PAM (groene doos) wordt gespleten met Cas9 complex en 3′ Da overhangen worden toegevoegd. B Adapters met 3 ‘ DT-overhang (blue box) zijn aan beide uiteinden van het decolletéproduct gelig., C Primers worden gebruikt om te verrijken voor PAM-zijdige gespleten producten door PCR., d Na PCR verrijking, DNA-fragmenten worden gezuiverd en Illumina compatibel ankers en barcodes zijn ‘staart-on’ door twee rondes van PCR (grijze vakken) en Illumina diep doorgeschoven
test moeten Cas9 PAM voorkeuren
De gerandomiseerde PAM bibliotheken beschreven in de vorige sectie werden onderworpen aan in-vitro digestie met verschillende concentraties van recombinant Cas9 eiwit geleverd met gids RNA om te gehalte Cas9 endonuclease PAM voorkeuren in een dosis-afhankelijke manier., Na de spijsvertering met Cas9-gidsrna ribonucleoprotein (RNP) complexen, werden de opeenvolgingscombinaties van PAM van de gerandomiseerde Pam bibliotheek die splitsing steunden gevangen door adapters aan de vrije einden van de plasmidedna molecules te ligeren die door het Cas9-gidsrna complex (Fig. 1a en b). Om efficiënte afbinding en vangst van de gespleten einden te bevorderen, werd de botte dubbelstrengelde die die DNA-snede door Cas9 endonucleases wordt geproduceerd gewijzigd om een 3′ Da-overhang te bevatten en werden de adapters gewijzigd om een aanvullende 3′ DT-overhang te bevatten., Om voldoende hoeveelheden DNA te produceren voor het rangschikken, werden de fragmenten van DNA die de PAM-opeenvolging Herbergen die splitsing ondersteunen PCR vergroot gebruikend een inleiding in de adapter en een ander direct naast het PAM-gebied (Fig. 1c). De resulterende PCR versterkte Cas9 PAM bibliotheken werden omgezet in ampli-seq templates (Fig. 1d) en single-read diep gerangschikt vanaf de adapter-kant van het amplicon., Om een adequate dekking te garanderen, werden de Cas9 PAM-bibliotheken gesequenced tot een diepte die minstens vijf keer groter was dan de diversiteit in de initiële gerandomiseerde PAM-bibliotheek (5.120 en 81.920 lezen voor respectievelijk de 5 en 7 gerandomiseerde BP PAM-bibliotheken). PAM sequenties werden geà dentificeerd uit de resulterende sequentiegegevens door alleen die reads te selecteren die een perfecte 12 nt sequentieovereenkomst bevatten die aan weerszijden van de 5 of 7 nt PAM sequentie flankeert (afhankelijk van de gebruikte gerandomiseerde PAM bibliotheek); alleen die PAM sequenties vastleggen die het resultaat zijn van perfecte Cas9-guide RNA target site herkenning en splitsing., Om de inherente bias in de initiële gerandomiseerde PAM bibliotheken te compenseren, werd de frequentie van elke PAM sequentie genormaliseerd naar zijn frequentie in de startbibliotheek. Aangezien de hier beschreven analyse Cas9-splitsbare Pam-sequenties direct vangt, werd probabilistische modellering gebruikt om de PAM-consensus voor elke Cas9-proteã ne te berekenen. Dit werd bereikt door de waarschijnlijkheid te evalueren van het vinden van elk nucleotide (G, C, A, of T) op elke positie van de Pam-sequentie onafhankelijk gebruikend een positiefrequentiematrix (PFM) (Methodensectie en )., De resulterende waarschijnlijkheden werden vervolgens gevisualiseerd als een WebLogo .
om de neiging tot vals-positieven in de test te onderzoeken, werd de toevoeging van Cas9 RNP-complexen in de digestiestap weggelaten (Fig. 1a) en de test werd uitgevoerd door de PCR-verrijkingsstap (Fig. 1c). Zoals getoond in aanvullend dossier 1: Figuur S5A, werden geen versterkingsproducten ontdekt in de absense van Cas9-gidsrna complexen. Dit betekent dat de incidentie van valse positieven laag is en niet significant bijdraagt aan de resultaten van de test.,
PAM-voorkeuren van Streptococcus pyogenes en Streptococcus thermophilus (CRISPR3-en CRISPR1-systemen) Cas9-eiwitten
om de test te valideren, werden de PAM-voorkeuren van Streptococcus pyogenes (Spy) en Streptococcus thermophilus CRISPR3 (Sth3) Cas9-eiwitten, waarvan de PAM-sequentiebehoefte eerder is gemeld , onderzocht. In vitro digests werden uitgevoerd met 1 µg (5,6 nM) van de 5 BP gerandomiseerde PAM bibliotheek bij twee concentraties, 0,5 en 50 nM, van voorgemonteerde spy of Sth3 Cas9 proteïne, crRNA, en tracrRNA RNP complexen gedurende 1 uur in een 100 µL reactievolume., Gebaseerd op hun frequentie in de 5 BP gerandomiseerde PAM bibliotheek, Spy en Sth3 Cas9 PAM sequenties (NGG en NGGNG, respectievelijk) waren bij definitieve concentraties van 0,40 nM en 0,11 nM in de spijsvertering, respectievelijk. Leden van de gerandomiseerde PAM bibliotheek die PAM sequenties bevatte die splitsing ondersteunden werden vastgelegd en geïdentificeerd zoals beschreven in de vorige sectie. Als negatieve controle, was de beginnende unclaved gerandomiseerde Pam bibliotheek onderworpen aan het rangschikken en PFM analyse naast die bibliotheken die aan Cas9 RNP complexen worden blootgesteld., Zoals getoond in aanvullend dossier 1: Figuur S5B en C, bestaan er geen opeenvolgingsvoorkeuren in de afwezigheid van Cas9 RNP complexe spijsvertering zoals duidelijk door een bijna perfecte verdeling van elk nucleotide op elke positie van PAM in de PFM-tabel en het gebrek aan informatieve inhoud in het WebLogo voor de controle. Dit is in grimmige beperkingen met Fig. 2a en b dat illustreert de samenstelling van de sequenties afgeleid van bibliotheken verteerd met Spy en Sth3 Cas9 RNP complexen. Onderzoek van de PFM afgeleide WebLogos (Fig., 2a en b) onthullen ook de aanwezigheid van de canonieke PAM voorkeuren voor de proteã nen van Spy en Sth3 Cas9, NGG en NGGNG , respectievelijk. Hoewel de PAM Voorkeuren gerapporteerd voor Spy en sth3 Cas9 proteã nen worden waargenomen in zowel de 0,5 nM als 50 nM verteert, is er een algemene verbreding in specificiteit onder de 50 nM verteert voorwaarden. Dit is het duidelijkst in positie 2 Voor het Spy Cas9 eiwit waar de frequentie van een niet-canoniek a residu dramatisch toeneemt (Fig. 2 bis)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
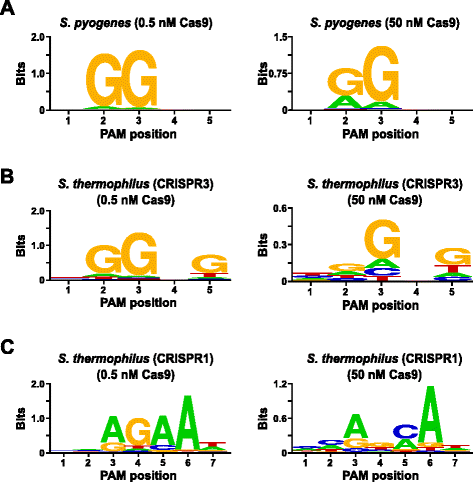
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., De frequentie van nucleotiden op elke PAM-positie werd onafhankelijk berekend met behulp van een position frequency matrix (PFM) en uitgezet als een WebLogo
verdere validatie van de test werd uitgevoerd door de PAM-voorkeuren voor het Streptococcus thermophilus CRISPR1 (Sth1) Cas9-eiwit te onderzoeken waarvan is gemeld dat de PAM-specificiteit zich uitstrekt tot 7 bp . Gebruikend 1 µg (5,6 nM) van de 7 BP gerandomiseerde PAM bibliotheek als malplaatje, werden de digesties van sth1 Cas9-gidsrna uitgevoerd bij twee concentraties, 0.5 nM en 50 nM, van complex RNP zoals hierboven beschreven., Als controles, Spy en Sth3 Cas9 RNP complexen werden ook gebruikt om de 7 BP gerandomiseerde PAM bibliotheek maar slechts bij enige, 0.5 nM, RNP complexe concentratie te verteren. Gebaseerd op de frequentie in de 7 BP gerandomiseerde PAM bibliotheek, waren de PAM sequenties eerder gerapporteerd voor Sth1 (NNAGAAW), Spy (NGG), en Sth3 (NGGNG) bij uiteindelijke concentraties van respectievelijk 0,01 nM, 0,22 nM en 0,05 nM., Zoals getoond in aanvullend dossier 1: Figuur S6A en B, waren de PAM voorkeuren voor Spy en sth3 Cas9 proteã nen die gebruikend de bibliotheek 7 BP worden geproduceerd bijna identiek aan die met de bibliotheek 5 BP worden geproduceerd die sterk bewijsmateriaal voor de reproduceerbaarheid van de analyse verstrekken. De PAM voorkeuren voor de sth1 Cas9 proteã ne kwamen ook nauw overeen die eerder, nnagaaw , bij de 0,5 nM Cas9-gids RNA complexe concentratie (Fig. 2c)., Gelijkaardig aan de Proteã nen van Spy en Sth3 Cas9, was Sth1 Cas9 in staat om een meer diverse reeks PAM opeenvolgingen in de reacties te splijten die een hogere concentratie van Cas9-gidsrna complex (50 nM) bevatten, was het opvallendste het duidelijke verlies van het residuvereiste van G op positie 4 en de bijna gelijke voorkeur voor een C en een bp op positie 5 (Fig. 2c). Dit resulteerde in een andere PAM-consensus dan die verkregen bij lagere concentraties.,
om te onderzoeken of PAM specificiteit onafhankelijk is van het type guide RNA, werden duplexed crRNA:tracrRNA of Sgrna , Spy, Sth3 en Sth1 Cas9 PAM Voorkeuren ook onderzocht met behulp van een binair Cas9 en sgRNA RNP complex. De spijsvertering werd uitgevoerd bij een enkele RNP complexe concentratie van 0,5 nM en de PAM voorkeurenanalyse werd uitgevoerd zoals hierboven beschreven. Zoals getoond in aanvullend dossier 1: Figuur S7A, B, en C, waren PAM Voorkeuren bijna identiek ongeacht het type van gebruikte gidsrna; of een crRNA: tracrRNA duplex of sgRNA., Bovendien, om te bevestigen dat PAM specificiteit niet sterk wordt beïnvloed door de samenstelling van de target DNA of spacer sequentie, werd de sequentie aan de andere kant van de 5 of 7 BP gerandomiseerde bibliotheek gericht op splitsing met een andere spacer; T2-5 (UCUAGAUAGAUUUACGAAUC) voor de 5 bp bibliotheek of T2-7 (CCGGCGACGUUGGGUCAACU) voor de 7 BP bibliotheek. De proteã nen van Spy en Sth3 Cas9 vooraf geladen met sgrnas die de T2 opeenvolging richten werden gebruikt om de 5 BP gerandomiseerde Pam bibliotheek te ondervragen terwijl de Sth1 Cas9-T2 sgrna complexen werden gebruikt om de 7 BP gerandomiseerde PAM bibliotheek te verteren., PAM voorkeuren werden beoordeeld zoals hierboven beschreven. De PAM voorkeuren voor alle 3 Cas9 proteã nen waren bijna identiek ongeacht de opeenvolging van spacer en doeldna (aanvullend dossier 1: Figuur S8A, B, en C).
Identificatie van sgrna-en PAM-voorkeuren voor het Brevibacillus laterosporus Cas9-eiwit
om de PAM-voorkeuren voor een Cas9-eiwit waarvan de PAM niet gedefinieerd was, empirisch te onderzoeken, werd een niet-gekarakteriseerde type II-C CRISPR-Cas-locus van de brevibacillus laterosporus-stam SSP360D4 (Blat) geïdentificeerd door interne DuPont Pioneer-databases voor Cas9-orthologen te doorzoeken., De locus (ongeveer 4.5 kb) bevatte een Cas9-gen dat een 1.092 polypeptide kon coderen, een CRISPR-array die zeven repeat-spacer-eenheden stroomafwaarts van het Cas9-gen en een tracrRNA-coderingsgebied stroomopwaarts van het Cas9-gen met gedeeltelijke homologie aan de CRISPR-array herhalingen omvat (Fig. 3a). De repeat en spacer Lengte (36 en 30 bp, dienovereenkomstig) is vergelijkbaar met andere type II CRISPR-Cas systemen met vijf van de acht herhalingen met 1 of 2 BP mutaties (Fig. 3b en aanvullend bestand 1: Figuur S9)., Andere genen typisch gevonden in een type II CRISPR-Cas locus waren of afgekapt (cas1) of ontbrekende (Fig. 3a).
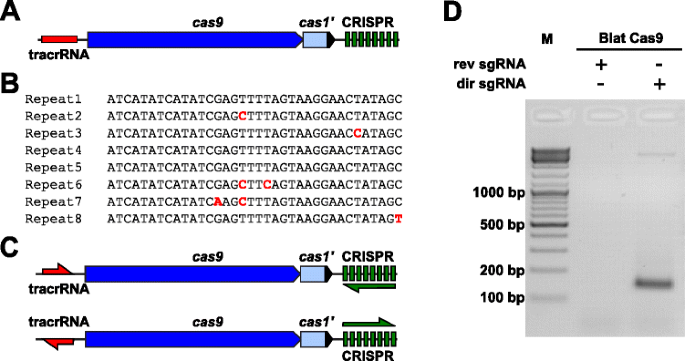
Identificatie van type II CRISPR-Cas-elementen in Brevibacillus laterosporus SSP360D4 CRISPR-Cas-systeem. een illustratie van het genomische DNA-gebied van het type II CRISPR-Cas-systeem van brevibacillus laterosporus SSP360D4. B vergelijking van type II CRISPR array repeat sequenties geïdentificeerd in Brevibacillus laterosporus SSP360D4., c de “direct” en “reverse” tracrrna en CRISPR array transcriptionele scenario’ s voor het type II CRISPR-Cas-systeem van Brevibacillus laterosporus SSP360D4. d Een agarosegel met reactieproducten, die aangeven dat alleen de ‘directe’ sgRNA (dir sgRNA), maar niet de ‘omgekeerde’ sgRNA (rev sgRNA) ondersteuning plasmide bibliotheek splijting in combinatie met de Cas9 endonuclease afkomstig van Brevibacillus laterosporus SSP360D4
De gids RNA-eis voor de Blat Cas9 eiwit werd bepaald door het genereren van twee sgRNA varianten., Deze varianten werden gegenereerd om zowel mogelijke zintuiglijke als anti-zintuiglijke uitdrukkingsscenario ‘ s van de reeks tracrRNA en CRISPR (Fig. 3c) en gebruikt om te onderzoeken welk expressiescenario splitsingsactiviteit van Blat Cas9 ondersteunde in de gerandomiseerde PAM-bibliotheek. De enige gids RNAs werd ontworpen door eerst de grenzen van de veronderstelde tracrrna molecules te identificeren door gebieden te analyseren die gedeeltelijk aan het 22 nt 5′ eindpunt van de herhaling (anti-herhaalt) waren., Vervolgens, om het 3′ eind van tracrRNA te bepalen, werden de mogelijke secundaire structuren en terminators gebruikt om het gebied van beëindiging in het stroomafwaartse fragment te voorspellen. Dit werd bereikt door het screenen op de aanwezigheid van Rho Onafhankelijke-als beëindigingsopeenvolgingen in het DNA rond de anti-herhaling gelijkend op die beschreven in Karvelis et al. , het omringende DNA omzetten in de opeenvolging van RNA en het onderzoeken van de resulterende structuren gebruikend UNAfold ., De resulterende sgRNAs werden ontworpen om een T7-het signaal van de initiatieherkenning van de polymerasetranscriptie aan het eind 5′ te bevatten gevolgd door een 20 nt doelherkenningsopeenvolging, 16 nt van crRNA-herhaling, 4 nt zelf-vouwende haarspeldlijn, en anti-herhaalopeenvolging complementair aan het herhaalde gebied van crRNA gevolgd door het resterende 3’ deel van vermeende tracrRNA. De sgrna-variant die een vermeende tracrRNA bevat die in dezelfde richting wordt getranscribeerd als het Cas9-gen (Fig. 3c) wordt ‘directe’ sgrna genoemd, terwijl de sgrna die de tracrRNA bevat in de tegenovergestelde richting een ‘omgekeerde’ sgRNA wordt getranscribeerd., Vijftig nM Van Blat Cas9 sgRNA RNP complex, vooraf geladen met respectievelijk de’ directe ‘of’ reverse ‘ sgRNAs, werden geïncubeerd met 1 µg (5,6 nM) van de 7 BP gerandomiseerde PAM bibliotheek. Na de spijsvertering van de bibliotheek en toevoeging van 3 ‘ Da overhangen, werden adapters gelig gemaakt en splitsingsproducten werden PCR versterkt (Fig. 1). Analyse van reactieproducten door agarose gelelektroforese toonde aan dat de’ directe ‘ sgrna, maar niet de ‘omgekeerde’ sgRNA plasmide bibliotheek splitsing ondersteund (Fig. 3d). De volgorde en de voorspelde secundaire structuur van de ‘directe’ sgrna zijn weergegeven in aanvullend bestand 1: Figuur S10.,
na het bepalen van de juiste guide RNA voor Blat Cas9, werd PAM identificatie uitgevoerd op dezelfde manier als hierboven beschreven voor de Spy, Sth3 en Sth1 Cas9 eiwitten tegen de 7 BP gerandomiseerde PAM bibliotheek met twee concentraties, 0,5 en 50 nM, van voorgemonteerde Blat Cas9 ‘direct’ sgrna RNP complex. Zoals in Fig. 4a, de PFM WEBLOGO PAM consensus voor het Blat Cas9 eiwit onder de 0,5 nM digest condities was NNNNCND (N = G, C, A, of T; D = A, G, of T) met een sterke voorkeur voor een C op positie 5 van de PAM sequentie., Een matige voorkeur voor een A werd waargenomen op positie 7 en lichte voorkeuren voor een C of T op positie 4 en G, C of a Boven T op positie 6 werden ook opgemerkt bij een zorgvuldige bestudering van de PFM-tabel (aanvullend dossier 1: Figuur S11). Evenzo aan Proteã nen van Spy, sth3, en Sth1 Cas9, verbreedt de PAM-specificiteit aangezien de complexe concentratie Cas9-sgRNA toeneemt. Dit is het duidelijkst op positie 5 waar een groter deel van PAM opeenvolgingen die een A residu steun splijten bij 50 nM in vergelijking met de 0.5 nM digest voorwaarden.
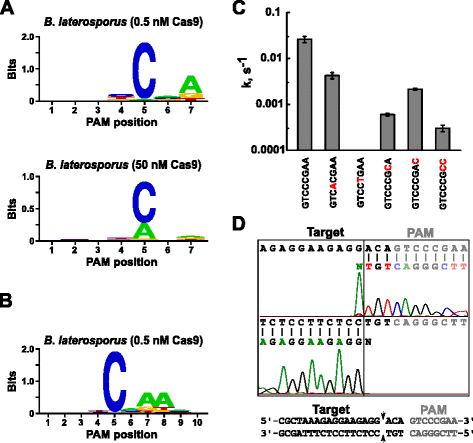
PAM voorkeuren en splitsingsposities van brevibacillus laterosporus SSP360D4 (Blat) Cas9 enzym. De voorkeuren van Blat Cas9 PAM toen 1 µg van bibliotheekdna met 0.5 nM of 50 Nm Cas9-sgrna complex (A) werd gespleten, breidde uit aan positie 10 door het protospacerdoel door 3 bp (b) te verschuiven. De frequentie van nucleotiden op elke PAM-positie werd onafhankelijk berekend met behulp van een position frequency matrix (PFM) en uitgezet als een WebLogo ., C Splitsingssnelheden van supercoiled plasmide DNA-substraten die mutaties bevatten (getoond in rood) in GTCCCGAA PAM sequentie. Alle gegevenspunten zijn gemiddelde waarden van ≥3 onafhankelijke experimenten. Foutbalken worden gegeven als S. D. d Run-off sequencing vanuit zowel de zintuiglijke als de anti-zintuiglijke richtingen van plasmide-DNA gespleten met Blat Cas9
aangezien Blat Cas9 elke base in de eerste drie posities van zijn PAM-sequentie kan accepteren (Fig. 4a), werd de afstandhouder T1 verschoven door drie nucleotiden in de richting 5′ om PAM-identificatie toe te staan die van 7 tot 10 bp moet worden uitgebreid., De verschoven T1 spacer, T1-3 (AAACGCUAAAGAGGAAGAGG), werd opgenomen in de Blat ‘directe’ sgrna en PAM identificatie werd uitgevoerd zoals eerder beschreven voor Spy, Sth3, Sth1 en Blat Cas9 eiwitten. De Pam-voorkeurenanalyse toonde aan dat de PAM-specificiteit voor Blat Cas9 kan worden uitgebreid tot positie 8 waar er een matige voorkeur is voor een extra A (Fig. 4b).
PAM-specificiteit voor Blat Cas9 werd bevestigd door plasmiden te genereren die mutaties bevatten in de meest geconserveerde residuen van de PAM (Fig. 4c)., De vervanging van het C-nucleotide op positie 5 schafte plasmidedna-splitsing af die zijn sleutelrol in Blat Cas9 PAM-erkenning bevestigt. Vervanging van een nucleotiden op posities 7 en 8 verminderde significant (respectievelijk 43× en 12×) de splitsingssnelheid van supercoiled plasmide die ook het belang van deze nucleotiden in Blat Cas9 PAM erkenning aangeeft.,
om de DNA-doelsplaatsen voor het Blat Cas9-eiwit te identificeren, werd een plasmide met een 20 bp-gebied dat overeenkomt met de spacer T1 gevolgd door een PAM-sequentie, GTCCCGAA, die binnen de PAM-consensus voor Blat Cas9, NNNNCNDD, viel, gegenereerd en verteerd met Blat Cas9-guide RNA-ribonucleoproteïnecomplex. Het directe rangschikken van DNA werd gebruikt om de einden van de lineaire molecule van DNA te bepalen die door Complex Blat Cas9 RNP wordt geproduceerd. De sequentieresultaten bevestigden dat plasmide DNA splitsing plaatsvond in de protospacer 3 nt 5 ‘ van de PAM sequentie (Fig., 4d) gelijkaardig aan dat waargenomen voor Spy, Sth3, en sth1 Cas9 proteã nen .
bij het bewerken van het planta-genoom met behulp van Blat Cas9 en sgRNA
na opheldering van de sgrna-en PAM-voorkeuren voor Blat Cas9, werden Cas9-en sgrna-expressiecassettes met geoptimaliseerde mais gegenereerd voor het testen van planta, zoals eerder beschreven voor het S. pyogenes cas9-gen en sgRNA . In het kort werd het Blat cas9-gen geoptimaliseerd voor maiscodon en intron 2 van het St-LSI-gen van de aardappel werd ingebracht om de expressie in E. coli te verstoren en optimale splicing in planta te vergemakkelijken (aanvullend dossier 1: Figuur S12)., De nucleaire lokalisatie van het Blat Cas9-eiwit in maïscellen werd vergemakkelijkt door de toevoeging van zowel amino-als carboxyl-terminale nucleaire locatiesignalen, SV40 (MAPKKKRKV) en Agrobacterium tumefaciens VirD2 (KRPRDRHDGELGGRKRAR), respectievelijk (aanvullend dossier 1: Figuur S12). Het gen Blat cas9 werd constitutief uitgedrukt in installatiecellen door geoptimaliseerde cas9 te koppelen aan een mais Ubiquitin promotor en pinII terminator in een plasmide DNA vector., Om een efficiënte sgrna-expressie in maïscellen te verkrijgen, werden een U6-polymerase III-promotor en-terminator (tttttttt) geïsoleerd en gefuseerd aan de 5′ – en 3′ – uiteinden van een gemodificeerde Blat sgRNA-coderingsvolgorde van DNA (aanvullend dossier 1: Figuur S13). De gewijzigde Blat sgRNA bevatte twee wijzigingen ten opzichte van die welke in de in vitro studies werden gebruikt; een T tot G-wijziging op positie 99 en een T tot C-wijziging op positie 157 van de sgRNA (aanvullend dossier 1: Figuur S13). De veranderingen werden geïntroduceerd om potentiële premature U6-polymerase III-terminatiesignalen in de Blat sgRNA te verwijderen., Wijzigingen werden aangebracht om een minimale impact te hebben op de secundaire structuur van de sgrna in vergelijking met de versie die wordt gebruikt in de in-vitro-studies (gegevens niet getoond).
om de mutatie-efficiëntie als gevolg van de onvolmaakte niet-homologe eindverbinding (nhej) reparatie van DNA-dubbelstrengbreuken (DSBS) als gevolg van spy en Blat Cas9-splitsing nauwkeurig te vergelijken, werden identieke genomische doelplaatsen van protospacer geselecteerd door doelen te identificeren met spy en Blat Cas9-compatibele PAMs, NGGNCNDD., Identieke spacer sequenties werden geselecteerd voor Blat en Spy Cas9 door het vastleggen van de 18 tot 21 nt sequentie direct stroomopwaarts van de PAM. Om optimale U6 polymerase III uitdrukking te verzekeren en geen mismatch binnen de sgrna spacer te introduceren, werden alle doelopeenvolgingen geselecteerd om op natuurlijke wijze in een G bij hun 5′ eind te beëindigen. Doelwitten werden geïdentificeerd en geselecteerd in exon 1 en 4 van het maisvruchtbaarheidsgen Ms45 en in een gebied stroomopwaarts van het mais liguleless-1-gen.,
De mutatieactiviteit van Blat Cas9 in maïs werd onderzocht door 10 dagen oude onvolgroeide maïsmembryo ‘ s (IMEs) biolistisch te transformeren met DNA-vectoren die cas9-en sgRNA-genen bevatten. Blat en de equivalente Spy Cas9-en sgRNA-expressievectoren werden onafhankelijk van elkaar in mais Hi-type II IMEs geïntroduceerd door middel van deeltjesverwerking zoals beschreven in . Omdat de transformatie van het deeltjespistool zeer variabel kan zijn, werd ook een visuele marker DNA-expressiecassette, DS-Red, samen met de Cas9-en sgRNA-expressievectoren geleverd om te helpen bij de selectie van gelijkmatig getransformeerde IMEs., In totaal werden drie transformatiereplicaten uitgevoerd op 60-90 IMEs en 20-30 van de meest gelijkmatig getransformeerde IMEs van elk replicaat werden 3 dagen na de transformatie geoogst. Het totale genomic DNA werd gehaald en het gebied dat de doelplaats omringt werd vergroot door PCR en amplicons die aan een gelezen diepte meer dan 300.000 worden gerangschikt. De resulterende reads werden onderzocht op de aanwezigheid van mutaties op de verwachte plaats van splitsing door vergelijking met controle-experimenten waar de sgrna DNA expressie cassette werd weggelaten uit de transformatie. Zoals in Fig., 5a, mutaties werden waargenomen op de verwachte plaats van splitsing voor Blat Cas9 met de meest voorkomende soorten mutaties die single base pair inserties of deleties. Soortgelijke reparatie patronen werden ook waargenomen voor de Spy Cas9 eiwit (extra bestand 1: Figuur S14 en). De mutatieactiviteit voor Blat Cas9 was robuust op twee van de drie geteste locaties en overtrof die van Spy Cas9 op de ms45 exon 4-doellocatie met ongeveer 30 % (Fig. 5b).
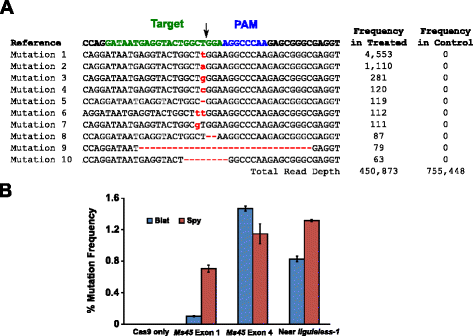
Brevibacillus laterosporus Cas9 bevordert nhej-mutaties in maïs. een Top 10 meest voorkomende soorten nhej mutaties gedetecteerd met Blat Cas9 in exon 4 van het ms45 gen. Een zwarte pijl geeft de verwachte plaats van splitsing aan; mutaties worden rood gemarkeerd; kleine letters geven een invoeging aan; ‘ – ‘ geeft een verwijdering aan. B vergelijking van Spy en Blat Cas9 nhej mutatiefrequenties op drie protospacer identieke doelplaatsen in maïs. Nhej mutaties werden gedetecteerd door deep sequencing 3 dagen na transformatie., Foutbalken vertegenwoordigen SEM, n = 3 deeltjespistool transformaties. Alleen Cas9 is de negatieve controle en vertegenwoordigt de gemiddelde achtergrondfrequentie (voor alle drie de doelplaatsen) van mutaties als gevolg van PCR-amplificatie en-sequencing