Embora voláteis compostos com atividade agonista foram analisadas em relação às Rup, um produto químico específico, com forte efeito agonista sobre mosquitos, 2-(4-Etil-5-(pyridin-3-il)-4H-1,2,4-triazole-3-ylthio)-N-(4-etilfenil)acetamida (VUAA1), abriu o campo de pesquisa (Jones et al., 2011; Taylor et al., 2012). Later studies by Taylor et al., (2012) forneceu elementos de prova de produtos químicos derivados da VUAA1, como a VUAA4, capazes de aumentar o seu efeito agonista em 10 vezes a ORco de A. gambiae, H. virescens e Harpegnathos saltator. Curiosamente, os autores relataram que qualquer mudança nos substituintes de amida causará uma completa perda de atividade agonista. Isto fornece informações úteis sobre os requisitos estruturais dos agonistas e a relação estrutura-atividade entre análogos e Rup da VUAA. Finalmente, apesar da actividade agonista potenciada dos produtos químicos da VUAA, o seu peso molecular relativamente elevado (367, 47 g mol−1 para a VUAA1) vs., agonistas voláteis, como bombykol (238,42 g mol−1), fazem com que uma entrega Volátil direta de VUAA algo não seja viável. Com isso em mente, a busca por análogos estruturais menores representa um foco interessante de pesquisa.antagonismo sobre a subunidade ORco
juntamente com o estudo de análogos relacionados com VUAA que podem atuar como agonistas fortes, o bloqueio de ORco pelos antagonistas também emergiu para guiar a concepção de semioquímicos e pesticidas., Assim, um estruturais analógico de VUAA1, VU0183254 (2-(4-Etil-5-furano-2-il-4H-triazole-3-ylsulfanyl)-1-fenotiazín-10-il-ethanone), foi relatado para inibir ORco resposta, atuando como alostéricos modulador em A. gambiae e perturbar o reconhecimento de agonistas como eugenol pelo complexo OR65/ORco (Jones et al., 2012). Outros análogos estruturais da VUAA também foram notificados como antagonistas. Um exemplo é o composto n -, 2-substituído de triazolotioacetamida OLC3 e OLC12 que interrompe a resposta ORco de uma forma semelhante em C. quinquefasciatus, A. gambiae, D. melanogaster, e O., nubilalis, sugerindo um local de ligação conservado em ORco (Chen e Luetje, 2012). Considerando a inibição do ORco como uma estratégia promissora para interromper comportamentos de insetos, parece que os esforços subsequentes devem apontar para os compostos com menor peso molecular do que antagonistas derivados de VUAA. Por exemplo, OX1a (232 g mol−1), triptaminas (160.22 g mol−1) e isopropílico cinnamate (190.24 g mol−1) foram relatados para ter efeito antagonista no ORco (Chen e Luetje, 2013, 2014; Tsitoura et al., 2015) com cerca de metade ou menos de peso molecular do que VUAA1., No entanto, o uso futuro destes antagonistas deve ser estudado cuidadosamente, uma vez que o bloqueio do ORco conservado pode afetar não só insetos prejudiciais, mas também benéficos.
além do efeito antagonista sondado in vitro, a evidência a nível comportamental suporta a ideia de que análogos estruturais das feromonas podem funcionar como antagonistas. Por exemplo, Sellanes et al., (2010) relataram a inibição da resposta sexual na honeydew traça Cryptoblabes gnidiella quando estruturais análogos, (Z)-9-tetradecenyl formiato de e (Z)-11-hexadecenyl formiato, foram adicionados sintético sexo feromônio, (Z)-11-hexadecenal e (Z)-13-octadecenal, em testes em túnel de vento. Este efeito antagonista da feromona foi posteriormente corroborado em ensaios de campo, onde a captura de machos da C. gnidiella diminuiu num padrão dose-dependente. O antagonismo das feromonas também foi relatado para B. mandarina, um ancestral de B. mori (Daimon et al., 2012)., Seus achados corroboram bombykol como o pheromone sexual, e bombykal e acetato de bombykyl como antagonistas, o que inibiu fortemente a atração de homens no campo para o pheromone bombykol sexual. Mais recentemente, evidências de antagonismo de feromonas foram relatadas para a mariposa do focinho Herpetogramma submarginale. Quando o (Z)-13-hexadecenol foi adicionado à sua feromona sexual, o acetato de (Z)-13-hexadecenilo diminuiu significativamente o número de machos capturados no campo (Yan et al., 2015)., O antagonismo das feromonas parece basear-se nas diferenças do grupo funcional químico, tais como álcoois, aldeídos e ésteres, dependendo da espécie de insecto. No entanto, o efeito antagonista destes análogos estruturais pode não ser devido à inibição ORco, mas à especificidade de ORx aos antagonistas. Um estudo recente sugere que o OR16 de Helicoverpa armigera é capaz de reconhecer especificamente o antagonista de feromonas, (Z)-11-hexadecenol (Chang H. et al., 2017). Os autores apoiaram o papel específico da OR16 considerando que H., as fêmeas de armigera emitem o composto antagonista juntamente com a sua feromona sexual (Z)-11-hexadecenal e(Z)-9-hexadecenal) como uma estratégia para evitar acasalamento Não-ideal com machos imaturos. Quando o OR16 foi derrubado pela técnica de edição do genoma CRISPR/Cas9 e os machos h. armigera foram testados por eletrofisiologia e testes comportamentais, nenhuma resposta da EAG foi registrada e os machos tentaram acasalar com fêmeas imaturas.
receptores Odorantes vs., Proteínas de ligação: prós e contras para o manejo de pragas de insetos
para o caso de OBPs, a especificidade ligando e mecanismos de OBPs representam aspectos controversos, que parecem fortemente dependentes dos métodos utilizados para a medição da afinidade ligando. Por exemplo, tem sido relatado que PBPs, tais como aqueles de P. xylostella das mariposas e eogystia hippophaecolus, podem ligar componentes de feromona sexual e análogos (Sun et al., 2013a; Hu et al., 2018). Isto sugere que jogadores a jusante, tais como ORs, poderiam aumentar a especificidade e a sensibilidade da recepção odorante., Evidências recentes sustentam que a co-expressão de PBPs e PRs pode aumentar a sensibilidade para feromonas. Por exemplo, múltiplas combinações de PR1-4 e PBP1-4 foram usadas para testar a sua resposta aos componentes de feromonas sexuais da traça Chilo supressalis (Chang et al., 2015). Os autores encontraram um aumento significativo na sensibilidade da resposta para (Z)-11-hexadecenal quando PR4 e PR6 foram co-expressos com PBP4., Embora a interação destas proteínas pode surgir um novo nível de investigação como o controle de pragas metas, os diferentes emparelhamento do PRs e Plps galpão luzes sobre a complexidade do sistema olfativo em insetos, fazendo com que a abordagem de uma tarefa difícil para um grande conjunto de compostos e de proteínas para teste. Apesar do acima exposto, os insectos OBP são de pequeno tamanho molecular, com fácil produção de proteínas recombinantes, o que os torna alvos favoritos para estudos estruturais e rastreio de ligação rápida., Por exemplo, a triagem de ligantes com OBPs poderia permitir a identificação de propriedades químicas para uma melhor ligação, tais como comprimento da cadeia, volume molecular, grupos funcionais e unsaturation da ligação. Estes, combinados com novos métodos de previsão de estruturas proteicas, como os utilizados no design de medicamentos e anticorpos médicos, como modelagem de homologia, simulações dinâmicas e acoplagem molecular, podem colocar OBPs de insetos em uma posição favorita sobre ORs como alvos para o desenvolvimento de agentes de controle na gestão de pragas.as ORs dos Insectos parecem mais especificamente adaptadas aos odores do que às OBPs., A maior especificidade demonstrada pelas ORs e a possibilidade de ativação/inibição de receptores específicos para um determinado comportamento tornam estas proteínas alvos atrativos para manipular comportamentos de pragas. A viabilidade da inibição do complexo ORx/ORco ou ORco por antagonistas inclui uma estratégia promissora para interromper o comportamento específico dos insetos, como o acasalamento através de receptores de feromonas sexuais. No entanto, a falta de informação estrutural é o ponto de estrangulamento na utilização de ORS de insectos como alvos para previsões de actividade semioquímica., As tabelas 1, 2 resumem o número de OBPs e ORs que foram identificados em espécies de insetos por transcriptoma (isto é, RNA-seq) e sequenciamento do genoma. A maioria dos insetos estudados até agora tem pelo menos duas vezes ORs do que OBPs de acordo com os estudos do genoma. Além disso, há uma extensa expansão de ORs em insetos sociais da ordem Hymenopterana, como a abelha melifera com 170 ORs (Weinstock et al., 2006), and the ants Solenopsis invicta and Cerapachys biroi with 400 and 506 ORs, respectively (Wurm et al., 2011; Oxley et al., 2014)., Da mesma forma, a expansão ou também é evidente em algumas pragas agrícolas, como o besouro da farinha vermelha T. castaneum com 265 ORs em comparação com 47 OBPs (Richards et al., 2008). Isso faz uma tarefa exigente para o alvo ou seleção, juntamente com a dificuldade para a expressão funcional de proteínas transmembranares, tais como ORs, a fim de rastrear um grande número de ligantes. Uma aproximação de propriedades importantes em ambos os OBPs e ORs são resumidas na Tabela 3.
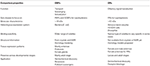
Quadro 3., Comparação aproximada de ORs de insectos e OBPs de acordo com as propriedades.
Outras Perspectivas
A caracterização funcional de insetos Rup, bem como a sua comprovada nas funções de insetos olfato lançar luzes sobre a sensibilidade e especificidade destes insetos proteínas específicas. Estes progressos reforçarão ainda mais a sua viabilidade como alvos de controlo de pragas, através da compreensão dos mecanismos de reconhecimento molecular e das interacções combinatórias com os OBPs., Por outro lado, o esforço massivo atual na identificação e caracterização de ligação de OBPs em várias espécies agrícolas importantes de insetos continuará e fornecerá mais informações sobre suas funções na fisiologia de insetos. Assim, esta revisão propõe como principal vantagem para OBPs sobre ORs, a disponibilidade de estruturas de cristal 3D e NMR, que com abordagens a jusante, como modelagem de homologia (quando necessário), acoplagem molecular e dinâmica molecular, iria refinar a busca de produtos químicos bioativos., Este último em complemento com a medição de afinidade ligando irá acelerar o estudo de OBPs de insetos a serem reconsiderados como os alvos para a descoberta semioquímica e as ferramentas para projetar super-ligandos no controle de pragas.a aparência e o desenvolvimento de resistência inseticida em pragas de insetos levaram à pesquisa intensiva sobre olfação de insetos e os mecanismos envolvidos no processamento neural. Está bem estabelecido que uma série de receptores e enzimas no SNC dos insectos são os alvos para o desenvolvimento de resistência aos insecticidas (Figura 1)., Foi demonstrado que a acetilcolinesterase (AChE) na forma solúvel proporciona resistência aos inseticidas organofosfórficos e carbamatos, agindo como bioscavengadores (Lee et al., 2015). Da mesma forma, foram demonstrados múltiplos mecanismos de Resistência a insecticidas no aphid M. persicae, envolvendo carboxilesterases, canais de sódio, ácido γ-aminobutírico (GABA) e nAChR (Bass et al., 2014)., Como componentes importantes na inseto periferia do sistema nervoso e os jogadores-chave no inseto comportamentos, tanto de insetos OBPs e Rup representam alvos alternativos para a identificação de compostos com semiochemical atividade (ou efeito agonista) e ferramentas para projeto forte de antagonistas para melhorar o desejado de respostas comportamentais de insetos pragas e reduzir o uso de inseticidas e subsequente resistência.
contribuições dos autores
HV escreveu secções sobre a estrutura e as RUP da OBP, desenvolveu tabelas e figura., J-JZ concebeu a ideia para o artigo de revisão, escreveu a seção principal, como introdução, função e estrutura OBPs, bem como a comparação ou-OBP.
Declaração de conflito de interesses
os autores declaram que a investigação foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.
agradecimentos
os autores gostariam de agradecer a FONDECYT 3170433., J-JZ está grato pelo apoio financeiro da Northeast Normal University e da Universidade Jilin, China por sua licença sabática para estudar na China. A pesquisa Rothamsted recebe uma bolsa com financiamento adicional de biotecnologia e Ciências Biológicas( BBSRC), Reino Unido.Gonzalez, F., Witzgall, P., and Walker, W. B. (2017). Transcriptomas antenais de três traças tortricidas revelam receptores quimiossensoriais putativos conservados para pistas olfativas e sociais. Ciência. Rep. 7, 1-12. doi: 10.,1038 / srep41829
PubMed Abstract | CrossRef Full Text/Google Scholar
Kaissling, K. E. (2013). A cinética das respostas olfativas pode depender em grande parte da interação odorante-receptor e da desativação odorante postulada para detectores de fluxo. J. Comp. Physiol. Neuroetol. Neural Sensorial Behav. Physiol. 199, 879–196. doi: 10.1007 / s00359-013-0812-z
PubMed Abstract / CrossRef Full Text | Google Scholar