o Projeto e construção de randomizados bibliotecas para análise a Cas9 PAM preferências
PAM bibliotecas que contém randomizados sequências de DNA imediatamente a jusante de uma sequência de DNA complementar para o espaçador de um guia de RNA foram gerados e utilizados para determinar empiricamente o PAM reconhecimento do Tipo II Cas9 endonucleases (Fig. 1)., Com a sequência-alvo do espaçador de ARN-guia sendo fixa, as bases randomizadas servem como um substrato para a leitura direta da especificidade de endonuclease PAM Cas9. Sequências aleatórias foram introduzidas em um vetor de DNA plasmídeo na região PAM de uma sequência alvo de protospacer demonstrando uma homologia perfeita para o espaçador de RNA guia T1 (CGCUAAAGAGAGAGAGAGAGAGGACA). Duas bibliotecas aumentando em tamanho e complexidade de cinco pares de base aleatórios (1.024 combinações PAM potenciais) para sete pares de base aleatórios (16.384 combinações PAM potenciais) foram geradas., Randomization of the 5 bp library was introduced through the synthesis of a single oligonucleotide containing five random residues. O oligonucleótido de cadeia simples foi convertido num modelo de cadeia dupla pela PCR (ficheiro adicional 1: Figura S1A), clonado no vector plasmídeo (ficheiro adicional 1: Figura S1B) e transformado em E. coli, tal como descrito na secção “métodos”., Para garantir a aleatoriedade ideal na biblioteca 7 bp PAM, o tamanho e a complexidade da biblioteca foram reduzidos pela síntese de quatro oligonucleótidos cada um contendo seis resíduos aleatórios mais um sétimo resíduo fixo compreendendo G, C, A ou T, respectivamente. Cada um dos quatro oligonucleótidos foi convertido separadamente em ADN de cadeia dupla, clonado em vector pTZ57R/T, tal como descrito na secção métodos, e transformado em E. coli, tal como descrito para a biblioteca 5 bp., Após a transformação, o DNA plasmídeo foi recuperado e combinado a partir de cada uma das quatro bibliotecas BP PAM 6 para gerar uma biblioteca BP PAM aleatória de 7 BP, compreendendo 16.384 possíveis combinações PAM. Para ambas as bibliotecas, a incorporação da aleatoriedade foi validada por sequenciação profunda; examinando a composição dos nucleótidos em cada posição da região PAM usando uma matriz de frequência de posição (PFM ) (seção de métodos e) (arquivo adicional 1: Figura S2A e B)., A distribuição e a frequência de cada sequência PAM na biblioteca PAM Aleatória 5 bp e 7 bp são mostradas no ficheiro adicional 1: figuras S3 e S4, respectivamente.
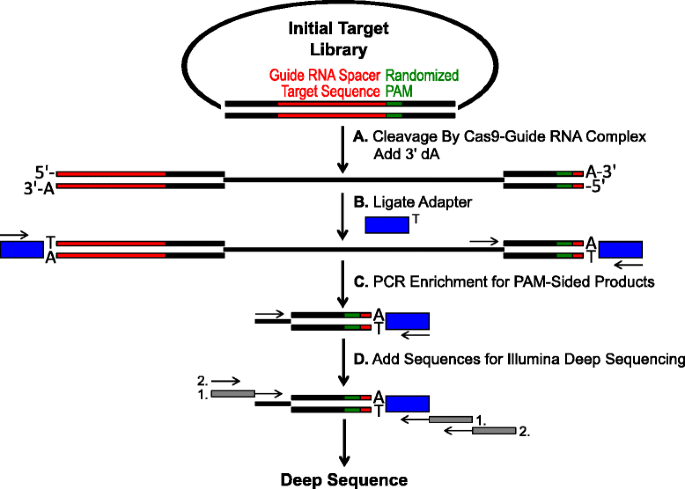
esquemático para identificação das preferências PAM pela clivagem Cas9 in vitro. uma biblioteca plasmídica inicial com PAM aleatória (caixa verde) é clivada com complexo Cas9 e 3′ dA overhangs são adicionados. Adaptadores b com overhang 3′ dT (blue box) são ligados a ambas as extremidades do produto de clivagem., C Primers são utilizados para enriquecer para produtos clivados de lado PAM por PCR., d Após a PCR enriquecimento de fragmentos de DNA são purificados e Illumina compatível âncoras e códigos de barras são ‘cauda-se através de duas rodadas de PCR (caixas cinzentas) e Illumina profunda seqüenciado
Análise a Cas9 PAM preferências
O randomizados PAM bibliotecas descrito na seção anterior, foram submetidas à digestão in vitro com diferentes concentrações de recombinante Cas9 proteína pré-carregado com guia de RNA, para ensaio Cas9 endonuclease PAM preferências de uma maneira dose dependente., Após digestão com complexos de ribonucleoproteína Rna-Guia Cas9 (RNP), combinações de sequências PAM da biblioteca Pam aleatorizada que suportava clivagem foram capturadas ligando Adaptadores às extremidades livres das moléculas de DNA plasmídeas clivadas pelo Complexo de RNA-guia Cas9 (Fig. 1a E b). Para promover a ligação eficiente e a captura das extremidades clivadas, o corte de DNA de ponta dupla gerado pelas endonucleases Cas9 foi modificado para conter um 3′ da overhang e Adaptadores foram modificados para conter um overhang 3′ DT complementar., Para gerar quantidades suficientes de ADN para sequenciamento, fragmentos de ADN que abrigavam a clivagem de suporte da sequência PAM foram amplificados pela PCR utilizando um iniciador no adaptador e outro directamente adjacente à região PAM (Fig. 1c). As bibliotecas amplificadas Cas9 PAM foram convertidas em modelos ampli-seq (Fig. 1d) e sequenciada a partir do adaptador-lado do amplificador., Para garantir uma cobertura adequada, as bibliotecas Cas9 PAM foram sequenciadas a uma profundidade pelo menos cinco vezes maior do que a diversidade na biblioteca inicial do PAM (5120 e 81.920 leituras para as bibliotecas aleatórias 5 e 7 bp PAM, respectivamente). Sequências PAM foram identificadas a partir dos dados de sequência resultantes, selecionando apenas as leituras que contêm uma combinação perfeita de 12 sequências nt flanqueando ambos os lados da seqüência 5 ou 7 nt PAM (dependendo da biblioteca Pam randomizada usada); capturando apenas as sequências PAM resultantes do reconhecimento e clivagem perfeito do local de destino de RNA Cas9-guia., Para compensar o viés inerente nas bibliotecas iniciais do PAM, a frequência de cada sequência do PAM foi normalizada para a sua frequência na biblioteca inicial. Uma vez que o ensaio aqui descrito captura directamente sequências PAM com clivagem Cas9, foi utilizada modelagem probabilística para calcular o consenso PAM para cada proteína Cas9. Isto foi realizado através da avaliação da probabilidade de encontrar cada nucleótido (G, C, A, ou T) em cada posição da sequência PAM de forma independente usando uma matriz de frequência de posição (PFM) (seção de métodos e )., As probabilidades resultantes foram então visualizadas como um WebLogo .
para examinar a propensão para falsos positivos no ensaio, a adição de complexos Cas9 RNP na fase de digestão foi omitida (Fig. 1a) e o ensaio foi realizado através da fase de enriquecimento de PCR(Fig. 1c). Como mostrado no arquivo adicional 1: Figura S5A, não foram detectados produtos de amplificação no absenso de complexos de RNA-guia Cas9. Assim, indicando que a incidência de falsos positivos é baixa e não contribui significativamente para os resultados do ensaio.,
PAM preferências de Streptococcus pyogenes e Streptococcus thermophilus (CRISPR3 e CRISPR1 sistemas) Cas9 proteínas
para validar o ensaio, o PAM preferências de Streptococcus pyogenes (Espião) e Streptococcus thermophilus CRISPR3 (Sth3) Cas9 proteínas, cuja PAM sequência de exigência tem sido relatado anteriormente , foram examinados. Foram realizadas digestões in vitro com 1 µg (5,6 nM) da biblioteca PAM aleatorizada de 5 bp em duas concentrações, 0,5 e 50 nM, de Espia pré-montada ou proteína Cas9 Sth3, crRNA, e complexos RNP de tracrRNA durante 1 h num volume de reacção de 100 µL., Com base na sua frequência na biblioteca PAM, Spy e sth3 Cas9 PAM (NGG e NGGNG, respectivamente) foram em concentrações finais de 0.40 nM e 0.11 nM na digestão, respectivamente. Membros da biblioteca Pam aleatorizada que continha sequências PAM que suportavam clivagem foram capturados e identificados como descrito na seção anterior. Como um controle negativo, A Biblioteca PAM inicial, não folheada, estava sujeita a sequenciação e análise PFM ao lado das bibliotecas expostas aos complexos Cas9 RNP., Como mostrado no arquivo Adicionais 1: Figura S5B e C, sem sequência preferências existir na ausência de Cas9 RNP complexo digestão como evidentes por uma perfeita distribuição de cada um dos nucleotídeos em cada posição do PAM na GFP quadro e a falta de conteúdo informativo no WebLogo para o controle. Isto é em stark construast com Fig. 2a e b que ilustra a composição das sequências derivadas de bibliotecas digeridas com complexos de Spy e Sth3 Cas9 RNP. Examination of the PFM derived WebLogos (Fig., 2a e b) também revelam a presença das preferências canônicas PAM para as proteínas Spy e Sth3 Cas9, NGG e NGGNG , respectivamente. Embora as preferências PAM relatadas para as proteínas Spy e Sth3 Cas9 sejam observadas tanto na digestão de 0,5 nM como 50 nM, há um alargamento geral na especificidade sob as condições de digestão de 50 nM. Isto é mais evidente na posição 2 para a proteína Espy Cas9, onde a frequência de um resíduo não canônico aumenta dramaticamente (Fig. 2a)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
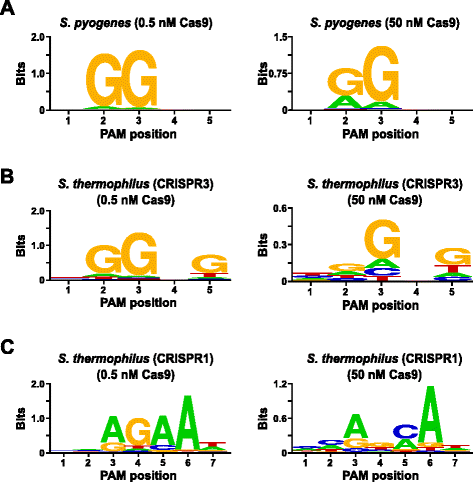
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., Frequência de nucleótidos em cada PAM posição foi de forma independente calculado utilizando-se uma posição de frequência matriz (GFP) e plotados como uma WebLogo
além disso, a validação do ensaio foi conduzido ao examinar o PAM preferências para o Streptococcus thermophilus CRISPR1 (Sth1) Cas9 proteína cuja PAM especificidade tem sido relatado para estender a 7 bp . Utilizando 1 µg (5,6 nM) da biblioteca PAM aleatorizada de 7 bp como modelo, as digestões de ARN-guia Cas9 do Sth1 foram realizadas em duas concentrações, 0,5 nM e 50 nM, do complexo RNP, como descrito acima., Como controles, Spy e sth3 complexos Cas9 RNP também foram usados para digerir a biblioteca PAM aleatorizada 7 bp, mas apenas em uma única concentração de 0,5 nM, complexo RNP. Com base na freqüência de 7 a bp randomizados biblioteca PAM, PAM sequências previamente descritos para Sth1 (NNAGAAW), Spy (NGG), e Sth3 (NGGNG) em concentrações finais de 0,01 nM, de 0,22 nM, e de 0,05 nM, respectivamente., Como mostrado no arquivo adicional 1: Figura S6A e B, as preferências PAM para proteínas Spy e Sth3 Cas9 geradas usando a biblioteca 7 bp foram quase idênticas às produzidas com a biblioteca 5 bp fornecendo fortes evidências para a reprodutibilidade do ensaio. As preferências PAM para a proteína Cas9 Sth1 também coincidiram estreitamente com as anteriormente relatadas, NNAGAAW, na concentração do complexo Cas9-guia RNA de 0, 5 nM (Fig. 2c)., Semelhante ao Espião e Sth3 Cas9 proteínas, Sth1 Cas9 foi capaz de romper mais um conjunto diversificado de PAM sequências de reações que contém uma maior concentração de Cas9-guia de RNA complexa (50 nM), a mais marcante foi a acentuada perda de G resíduo de exigência na posição 4 e perto de igual preferência por um C e Um bp na posição 5 (Fig. 2c). Isto resultou num consenso PAM diferente do obtido em concentrações mais baixas.,
Para examinar se PAM especificidade é independente do tipo de guia de RNA, duplex crRNA:tracrRNA ou sgRNA , Espião, Sth3, e Sth1 Cas9 PAM preferências também foram examinados usando um binário Cas9 e sgRNA RNP complexo. A digestão foi realizada com uma única concentração de complexo RNP de 0, 5 nM e a análise de preferência PAM foi realizada como descrito acima. Como mostrado no arquivo adicional 1: Figura S7A, B, E C, as preferências PAM eram quase idênticas independentemente do tipo de RNA guia utilizado; ou um crRNA:tracrRNA duplex ou sgRNA., Além disso, para confirmar que o PAM especificidade não é muito influenciado pela composição do DNA alvo ou espaçador seqüência, a seqüência, no lado oposto de 5 ou 7 bp randomizados biblioteca foi alvo de clivagem com diferentes espaçadores; T2-5 (UCUAGAUAGAUUACGAAUUC) 5 bp de biblioteca ou T2-7 (CCGGCGACGUUGGGUCAACU) para o 7 bp biblioteca. Spy e Sth3 Cas9 proteínas pré-carregado com sgRNAs segmentação T2 sequência foram utilizados para interrogar a 5 bp randomizados biblioteca PAM enquanto o Sth1 Cas9-T2 sgRNA complexos foram utilizados para digerir a 7 bp randomizados biblioteca PAM., As preferências da PAM foram avaliadas como descrito acima. As preferências PAM para todas as 3 proteínas Cas9 foram quase idênticas independentemente da sequência de ADN-alvo e espaçador (ficheiro adicional 1: Figura S8A, B E C).
a Identificação de sgRNA e PAM preferências para o Brevibacillus laterosporus Cas9 proteína
Para examinar empiricamente o PAM preferências para uma Cas9 proteína cuja PAM estava indefinido, um descaracterizada Tipo II-C CRISPR-Cas locus de Brevibacillus laterosporus tensão SSP360D4 (Blat) foi identificado através de pesquisa interna DuPont Pioneer bancos de dados para Cas9 orthologues., O locus (aproximadamente 4,5 kb) continha uma cas9 gene capaz de codificação em 1.092 um polipeptídeo, um CRISPR matriz, formada por sete repita-espaçador de unidades a jusante do cas9 gene e um tracrRNA codificação de região localizada a montante do cas9 gene com parciais de homologia para a CRISPR matriz repete-se (Fig. 3a). O comprimento de repetição e espaçador (36 e 30 bp, em conformidade) é semelhante a outros sistemas CRISPR-Cas tipo II, com cinco das oito repetições contendo 1 ou 2 mutações bp (Fig. 3B e Ficheiro adicional 1: Figura S9)., Outros genes tipicamente encontrados em um locus tipo II CRISPR-Cas foram truncados (cas1) ou ausentes (Fig. 3a).
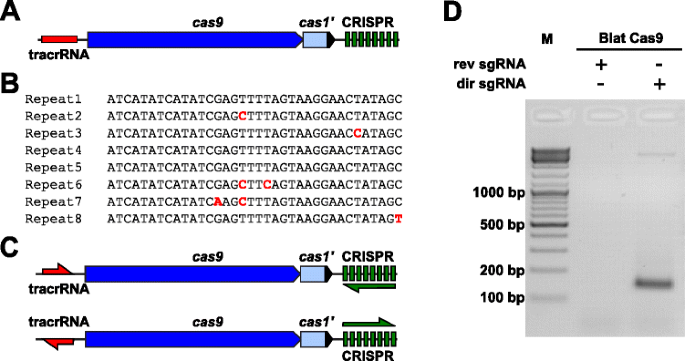
a Identificação do Tipo II CRISPR-Cas elementos Brevibacillus laterosporus SSP360D4 CRISPR-sistema Cas. a ilustration of the genomic DNA region from the Type II CRISPR-Cas system from Brevibacillus laterosporus SSP360D4. b comparação da matriz CRISPR tipo II sequências repetidas identificadas em Brevibacillus laterosporus SSP360D4., c os cenários de transcrição “directos” e “inversos” do tracrRNA e do CRISPR para o sistema CRISPR-Cas tipo II do Brevibacillus laterosporus SSP360D4. d Um gel de agarose com produtos de reação, indicando que somente o ‘direto’ sgRNA (dir sgRNA), mas não o ‘inverso’ sgRNA (rev sgRNA) apoio plasmídeo biblioteca de clivagem em combinação com a Cas9 endonuclease provenientes de Brevibacillus laterosporus SSP360D4
A guia de RNA requisito para o Blat Cas9 de proteína foi determinada por gerar dois sgRNA variantes., Estas variantes foram geradas para explicar tanto os possíveis cenários de expressão sensorial ou anti-sensorial do tracrRNA e array CRISPR(Fig. 3c) e usado para sondar qual cenário de expressão suportava a atividade de clivagem de Blat Cas9 na biblioteca Pam aleatória. O Guia Único RNAs foi projetado pela primeira vez identificando os limites das moléculas putativas tracrRNA através da análise de regiões que eram parcialmente complementares ao terminus 22 nt 5′ da repetição (anti-repetição)., Em seguida, para determinar a extremidade 3′ do tracrRNA, possíveis estruturas secundárias e terminadores foram usados para prever a região de terminação no fragmento a jusante. Isto foi realizado através da triagem para a presença de sequências de terminação Tipo Rho independente no DNA em torno da anti-repetição semelhante ao descrito em Karvelis et al. , convertendo o DNA circundante em sequência de RNA e examinando as estruturas resultantes usando UNAfold ., As sgRNAs resultantes foram projetadas para conter um sinal de reconhecimento de iniciação da transcrição polimerase T7 no final de 5′, seguido por uma sequência de reconhecimento de alvo de 20 nt, 16 nt de repetição crRNA, 4 nt de espiral auto-dobrável, e sequência anti-repetição complementar à região de repetição do crRNA seguido pela parte restante de 3′ da tracrRNA putativa. A variante sgRNA que contém um putativo tracrRNA transcrito na mesma direção que o gene cas9 (Fig. 3c) é denominado sgRNA ‘direto’, enquanto o sgRNA contendo o tracrRNA transcrito na direção oposta um sgRNA ‘reverso’., 50 nM de Complexo Blat Cas9 sgRNA RNP, pré-carregado com o sgRNAs “direto” ou “reverso”, respetivamente, foram incubados com 1 µg (5,6 nM) da biblioteca PAM aleatorizada de 7 bp. Após a digestão da biblioteca e a adição de 3 ‘ dA overhangs, adaptadores foram ligados e produtos de clivagem foram amplificados PCR (Fig. 1). A análise dos produtos de reacção pela electroforese em gel de agarose revelou que o sgRNA “directo”, mas não o sgRNA “reverso”, suportava a clivagem da biblioteca plasmídica (Fig. 3d). A sequência e a estrutura secundária prevista do sgRNA “directo” são apresentadas no ficheiro adicional 1: Figura S10.,
Após a determinação do guia de RNA para Blat Cas9, PAM identificação foi realizada de forma semelhante ao descrito acima para o Espião, Sth3, e Sth1 Cas9 proteínas contra o 7 bp randomizados biblioteca PAM com duas concentrações de 0,5 e 50 nM, de pré-montados Blat Cas9 ‘direto’ sgRNA RNP complexo. Como mostrado na Fig. 4a, o PFM WebLogo PAM consensus for the Blat Cas9 protein under the 0.5 nM digest conditions was nnncnd (N = G, C, A, or T; D = A, G, or T) with a strong preference for a C at position 5 of the PAM sequence., Observou-se também uma preferência moderada por um A na posição 7 e preferências ligeiras para um C ou T na posição 4 E G, C, ou a sobre T na posição 6 ao examinar atentamente a tabela PFM (ficheiro adicional 1: Figura S11). Similarmente às proteínas Spy, Sth3 e Sth1 Cas9, a especificidade PAM amplia-se à medida que a concentração do complexo Cas9-sgRNA aumenta. Isto é mais evidente na posição 5, onde uma maior proporção de sequências PAM contendo uma clivagem de suporte de resíduos a 50 nM, em comparação com as condições de digestão de 0,5 nM.
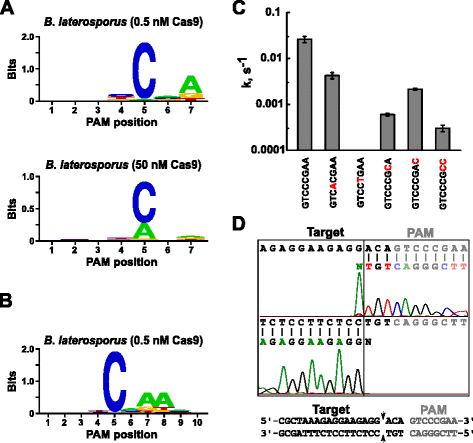
PAM preferências e clivagem posições de Brevibacillus laterosporus SSP360D4 (Blat) Cas9 enzima. Blat Cas9 PAM preferences when 1 µg of library DNA was cleaved with 0.5 nM or 50 nM Cas9-sgRNA complex (a), extended out to position 10 by shifting the protospacer target by 3 bp (b). A frequência de nucleótidos em cada posição PAM foi calculada independentemente usando uma matriz de frequência de posição (PFM) e plotada como um WebLogo ., c taxas de clivagem de substratos de ADN plasmídeo supercoolados contendo mutações (apresentadas a vermelho) na sequência PAM da GTCCGAA. Todos os pontos de dados são valores médios de ≥3 experiências independentes. As barras de erro são dadas como S. D. d de Run-off de seqüenciamento de tanto senso e anti-senso direções de plasmídeo de DNA clivado com Blat Cas9
Desde Blat Cas9 pode aceitar qualquer base nas três primeiras posições dos seus PAM sequência (Fig. 4a), o espaçador T1 foi deslocado por três nucleotídeos na direção 5′ para permitir que a identificação PAM fosse estendida de 7 para 10 bp., O espaçador T1 deslocado, T1-3 (AAACGCUAAAGAGAGAGG), foi incorporado ao blat ‘direct’ sgRNA e a identificação PAM foi realizada como descrito anteriormente para as proteínas Spy, Sth3, Sth1 e Blat Cas9. A análise de preferência PAM revelou que a especificidade PAM para Blat Cas9 pode ser estendida para a posição 8, onde existe uma preferência moderada por um A adicional(Fig. 4b).
PAM a especificidade para Blat Cas9 foi confirmada pela geração de plasmídeos como contendo mutações nos resíduos mais conservados do PAM(Fig. 4c)., A substituição do nucleótido C na posição 5 aboliu a clivagem do ADN plasmídeo confirmando o seu papel fundamental no reconhecimento do Blat Cas9 PAM. A substituição de um nucleótido nas posições 7 e 8 reduziu significativamente (43× e 12×, respectivamente) a taxa de clivagem do plasmídeo supercoilado também indicando a importância destes nucleótidos no reconhecimento Blat Cas9 PAM.,
para identificar as posições de clivagem do alvo de ADN para a proteína Blat Cas9, um plasmídeo contendo uma região de 20 bp correspondente ao espaçador T1 seguido de uma sequência PAM, GTCCGAA, abrangida pelo consenso PAM para Blat Cas9, NNNNCNDD, foi gerado e digerido com complexo de ribonucleoproteína RNA-guia Blat Cas9. A sequenciação direta do DNA foi usada para determinar as extremidades da molécula linear de DNA gerada pelo Complexo Blat Cas9 RNP. Os resultados da sequência confirmaram que a clivagem do ADN plasmídeo ocorreu no protospacer 3 nt 5′ da sequência PAM(Fig., 4d) similar ao observado para as proteínas Spy, Sth3 e Sth1 Cas9 .
na edição do genoma da planta utilizando Blat Cas9 e sgRNA
após elucidação das preferências de sgRNA e PAM para o Blat Cas9, foram geradas cassetes de expressão Cas9 e sgRNA optimizadas para os testes de planta, tal como descrito anteriormente para os genes cas9 e sgRNA de S. pyogenes . Brevemente, o gene Blat cas9 foi o codon de milho optimizado e o intron 2 do gene St-LSI da batata foi inserido para perturbar a expressão na E. coli e facilitar a articulação óptima no planta (ficheiro adicional 1: Figura S12)., A localização Nuclear da proteína Blat Cas9 nas células de milho foi facilitada pela adição de sinais de localizações nucleares amino e carboxil-terminais, SV40 (MAPKKKRKV) e Agrobacterium tumefaciens VirD2 (KRPRDRHDGELGGRKRAR), respectivamente (ficheiro adicional 1: Figura S12). O gene Blat cas9 foi constitucionalmente expresso em células vegetais, ligando o cas9 optimizado a um promotor de ubiquitina de milho e ao terminador pinII num vector de ADN plasmídeo., Para conferir uma expressão eficaz do sgRNA nas células de milho, um promotor e terminador de polimerase III (TTTTTTTTTT) de milho U6 foram isolados e fundidos às extremidades de 5′ e 3′ de uma sequência de ADN codificada do Blat sgRNA modificada, respectivamente (ficheiro adicional 1: Figura S13). O Blat sgRNA modificado continha duas modificações a partir das utilizadas nos estudos in vitro: uma alteração T A G na posição 99 e uma modificação T A C na posição 157 do sgRNA (ficheiro adicional 1: Figura S13). As alterações foram introduzidas para remover potenciais sinais prematuros de terminação da U6 polimerase III no blat sgRNA., Alterações introduzidas que tenham um impacto mínimo na estrutura secundária do sgRNA em comparação com a versão utilizada nos estudos in vitro (dados não apresentados).
para comparar com precisão a eficiência mutacional resultante da junção final não homóloga imperfeita (NHEJ) da reparação de quebras de dupla cadeia de ADN (DSBs) resultantes da clivagem Spy e Blat Cas9, os sítios-alvo genómicos idênticos protospacer foram seleccionados através da identificação de alvos com spy e Blat Cas9 compatíveis, NGGNCNDD., Sequências Spacer idênticas foram selecionadas para Blat e Spy Cas9 capturando a sequência 18 a 21 nt imediatamente a montante do PAM. Para garantir uma expressão óptima de U6 polimerase III e não introduzir um desfasamento dentro do espaçador sgRNA, todas as sequências-alvo foram seleccionadas para terminar naturalmente num G no seu fim de 5′. Os alvos foram identificados e seleccionados no exon 1 e 4 do gene de fertilidade do milho Ms45 e numa região a montante do gene do milho sem Ligadura-1.,a actividade mutacional do Blat Cas9 no milho foi examinada através da transformação biolística de embriões de milho imaturo de 10 dias com vectores de ADN contendo genes cas9 e sgRNA. Os vetores de expressão “Blat” e os vetores de expressão “Spy Cas9” e “sgRNA” equivalentes foram introduzidos de forma independente em IMEs de milho Hi-Tipo II por transformação de pistolas de partículas semelhantes às descritas em . Uma vez que a transformação de armas de partículas pode ser altamente variável, um cassette de expressão de ADN marcador visual, Ds-Red, também foi co-entregue com os vetores de expressão Cas9 e sgRNA para ajudar na seleção de IMEs uniformemente transformados., No total, três replicados de transformação foram realizados em 60-90 IMEs e 20-30 dos IMEs mais uniformemente transformados de cada replicado foram colhidos 3 dias após a transformação. O DNA genômico Total foi extraído e a região em torno do local alvo foi amplificada por PCR e amplificadores sequenciados a uma profundidade de leitura superior a 300.000. The resulting reads were examined for the presence of mutations at the expected site of clivage by comparison to control experiments where the sgRNA DNA expression cassette was omitted from the transformation. Como mostrado na Fig., 5a, foram observadas mutações no local esperado de clivagem para Blat Cas9 com os tipos mais prevalentes de mutações sendo inserções ou deleções de pares de base únicos. Padrões similares de reparo também foram observados para a proteína Spy Cas9 (arquivo adicional 1: Figura S14 e ). A atividade mutacional para Blat Cas9 foi robusta em dois dos três locais testados e excedeu a do Spy Cas9 no Ms45 exon 4 em aproximadamente 30% (Fig. 5b).
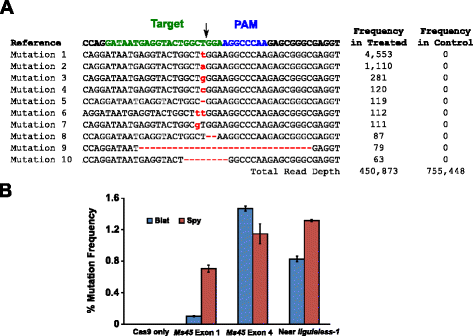
Brevibacillus laterosporus Cas9 promove NHEJ mutações do milho. um dos 10 tipos mais prevalentes de mutações NHEJ detectadas com Blat Cas9 no exon 4 do gene Ms45. Uma seta preta indica o local esperado de clivagem; as mutações são realçadas a vermelho; a letra de minúscula indica uma inserção; o ‘-‘ indica uma remoção. b Comparação das frequências de mutação Spy e Blat Cas9 NHEJ em três locais alvo idênticos em protospacer no milho. As mutações NHEJ foram detectadas por sequenciação profunda 3 dias após a transformação., As barras de erro representam transformações SEM, n = 3 de armas de partículas. Cas9 é apenas o controlo negativo e representa a frequência de fundo média (nos três locais-alvo) das mutações resultantes da amplificação e sequenciação da PCR