Deși volatile compusi cu activitate agonistă au fost verificate împotriva ORs, anumite substanțe chimice, cu puternic efect agonist pe tantari, 2-(4-Etil-5-(piridin-3-yl)-4H-1,2,4-triazol-3-ylthio)-N-(4-ethylphenyl)acetamidă (VUAA1), a deschis domeniul de cercetare (Jones et al., 2011; Taylor și colab., 2012). Studii ulterioare de Taylor și colab., (2012) a furnizat dovezi de VUAA1 derivate din substanțe chimice, cum ar fi VUAA4, capabil să-și mărească efect agonist de 10 ori pe ORco de A. gambiae, H. virescens, și Harpegnathos saltator. Interesant, autorii au raportat că orice modificare a substituenților de amidă va provoca o pierdere completă a activității agoniste. Acest lucru oferă informații utile despre cerințele structurale ale agoniștilor și relația structură-activitate dintre analogii VUAA și ORs. În cele din urmă, în ciuda îmbunătățită agonist activitatea de VUAA chimice, relativ înaltă greutate moleculară (367.47 g mol−1 pentru VUAA1) vs, volatile agoniști, cum ar fi bombykol (238.42 g mol−1), face o directă volatile livrare de VUAA ceva nu este fezabil. Având în vedere acest lucru, căutarea analogilor structurali mai mici reprezintă un obiectiv interesant al cercetării.antagonismul pe subunitatea ORco împreună cu studiul analogilor legați de VUAA care pot acționa ca agoniști puternici, blocarea ORco de către antagoniști a apărut, de asemenea, pentru a ghida semiochimicele și designul pesticidelor., Astfel, un analog structural al VUAA1, VU0183254 (2-(4-Etil-5-furan-2-il-4H-triazol-3-ylsulfanyl)-1-fenotiazinil-10-il-ethanone), a fost raportat de a inhiba ORco răspuns, care acționează ca modulator alosteric în A. gambiae și perturbă recunoașterea de agoniști, cum ar fi eugenolul de complex OR65/ORco (Jones et al., 2012). Alți analogi structurali VUAA au fost, de asemenea, raportați ca antagoniști. Un exemplu este N-,2-substituiți triazolothioacetamide compuși OLC3 și OLC12 care perturba ORco răspuns într-o manieră similară în C. quinquefasciatus, A. gambiae, D. melanogaster, și O., nubilalis, sugerând un sit de legare conservat în ORco (Chen și Luetje, 2012). Având în vedere inhibarea ORco ca o strategie promițătoare de a perturba comportamentele insectelor, se pare că eforturile ulterioare ar trebui să vizeze compușii cu greutate moleculară mai mică decât antagoniștii derivați de VUAA. De exemplu, OX1a (232 g mol−1), triptamină (160.22 g mol−1) și izopropilic cinamat (190.24 g mol−1) au fost raportate au efect antagonist pe ORco (Chen și Luetje, 2013, 2014; Tsitoura et al., 2015) cu aproximativ jumătate sau mai puțin greutate moleculară decât VUAA1., Cu toate acestea, utilizarea viitoare a acestor antagoniști ar trebui studiată cu atenție, deoarece blocarea ORco conservată poate afecta nu numai insectele dăunătoare, ci și cele benefice.pe lângă efectul antagonist probat in vitro, dovezile la nivel comportamental susțin ideea că analogii structurali ai feromonilor pot funcționa ca antagoniști. De exemplu, Sellanes și colab., (2010) au raportat inhibarea răspunsului sexual în mană molie Cryptoblabes gnidiella când analogii structurale, (Z)-9-tetradecenyl formiat și (Z)-11-hexadecenyl formiat, au fost adăugate la sex sintetic de feromoni, (Z)-11-hexadecenal și (Z)-13-octadecenal, în teste în tunelul aerodinamic. Acest efect antagonist de feromoni a fost ulterior coroborat în testele de teren, unde prinderea masculilor C. gnidiella a scăzut într-un model dependent de doză. Antagonismul feromonilor a fost, de asemenea, raportat pentru B. mandarina, un strămoș al lui B. mori (Daimon et al., 2012)., Concluziile lor confirma bombykol ca sexul feromoni, și bombykal și bombykyl acetat de ca antagoniști, care puternic inhibată atragerea masculilor în domeniul sex-feromon bombykol. Mai recent, dovezi de feromoni antagonism a fost raportat pentru bot molie Herpetogramma submarginale. Când s-a adăugat (Z)-13-hexadecenol la feromonul său sexual, (Z)-13-acetat de hexadecenil, a scăzut semnificativ numărul de bărbați capturați în câmp (Yan et al., 2015)., Antagonismul feromonilor pare bazat pe diferențele dintre grupurile funcționale chimice, cum ar fi alcoolii, aldehidele și esterii, în funcție de speciile de insecte. Cu toate acestea, efectul antagonist al acestor analogi structurali s-ar putea să nu se datoreze inhibării ORco, ci specificității ORx la antagoniști. Un studiu recent sugerează că OR16 de Helicoverpa armigera este capabil să recunoască în mod specific antagonist de feromoni, (Z)-11-hexadecenol (Chang H. et al., 2017). Autorii au susținut rolul specific al OR16, considerând că H., armigera femelele emit antagonist compus împreună cu feromoni sex ((Z)-11-hexadecenal și (Z)-9-hexadecenal) ca o strategie pentru a evita non-optime împerechează cu masculii imaturi. În mod remarcabil, când OR16 a fost doborât de tehnica de editare a genomului CRISPR / Cas9 și bărbații H. armigera au fost testați prin electrofiziologie și teste comportamentale, nu a fost înregistrat niciun răspuns EAG și bărbații au încercat să se împerecheze cu femele imature.
Receptorii odoranți vs., Proteine de legare: argumente Pro și Contra pentru Insecte Pest Management
Pentru cazul OBPs, ligand specificul și mecanismele de OBPs reprezintă aspecte controversate, care pare a fi puternic dependentă de metodele utilizate pentru măsurarea ligand de afinitate. De exemplu, s-a raportat că leagă penicilinele, cum ar fi cele de la molii P. xylostella și Eogystia hippophaecolus, se poate lega atât de sex feromoni componente și analogii (Sun et al., 2013a; Hu și colab., 2018). Acest lucru sugerează că jucătorii din aval, cum ar fi ORs, ar putea spori specificitatea și sensibilitatea recepției odorante., Dovezi recente susține că co-exprimare a PBPs și PRs poate crește sensibilitatea față de feromoni. De exemplu, mai multe combinații de PR1-4 și PBP1-4 au fost folosite pentru a testa reactia lor la sex feromoni componente ale molie Chilo suppressalis (Chang et al., 2015). Autorii au descoperit o creștere semnificativă a sensibilității răspunsului la (Z)-11-hexadecenal atunci când PR4 și PR6 au fost co-exprimate cu PBP4., Deși interacțiunea acestor proteine ar putea apărea un nou nivel de cercetare ca ținte de combatere a dăunătorilor, asocierea diferită a PRs și PBPs aruncă lumini asupra complexității sistemului olfactiv la insecte, făcând abordarea o sarcină dificilă pentru un set mare de compuși și proteine de testat. În ciuda celor de mai sus, OBP-urile insectelor sunt de dimensiuni moleculare mici, cu o producție ușoară de proteine recombinante, ceea ce le face ținte preferate pentru studiile structurale și screeningul rapid de legare., De exemplu, ligand de screening cu OBPs ar putea permite identificarea unor proprietăți chimice pentru o mai bună legare, cum ar fi lungimea lanțului molecular volum, grupe funcționale, și obligațiuni de nesaturare. Acestea, combinate cu noua structura de proteine metode de predicție cum este utilizat în proiectarea de medicamente medicale și anticorpi, cum ar fi omologie de modelare, dinamica simulări, și molecular docking, ar putea plasa insecte OBPs într-o poziție preferată de peste ORs ca ținte pentru dezvoltarea de agenți de control în gestionarea dăunătorilor.
ORS insecte par mai precis reglate la odoranți decât OBPs., Specificitatea mai mare arătată de ORs și șansa de activare/inhibare a receptorilor specifici pentru un anumit comportament fac ca aceste proteine să fie ținte atractive pentru manipularea comportamentelor dăunătorilor. Fezabilitatea inhibării complexului ORx/ORco sau ORco de către antagoniști cuprinde o strategie promițătoare de a perturba comportamentul specific al insectelor, cum ar fi împerecherea prin intermediul receptorilor de feromoni sexuali. Cu toate acestea, lipsa informațiilor structurale este blocajul în utilizarea rup a insectelor ca ținte pentru predicțiile activității semiochimice., Tabelele 1, 2 rezumă numărul de OBPs și rup care au fost identificate la speciile de insecte prin transcriptom (adică ARN-seq) și secvențierea genomului. Majoritatea insectelor studiate până în prezent au cel puțin de două ori ORs decât OBPs, conform studiilor genomului. Mai mult, există o extindere extinsă a ORs în insectele sociale din ordinul Hymenopteran, cum ar fi albina de miere A. mellifera cu 170 ORs (Weinstock et al., 2006), și furnicile Solenopsis invicta și Cerapachys biroi cu 400 și 506 ORs, respectiv (Wurm et al., 2011; Oxley și colab., 2014)., În mod similar, extinderea OR este evidentă și la unii dăunători agricoli, cum ar fi gândacul de făină roșie T. castaneum cu 265 ORs comparativ cu 47 OBPs (Richards et al., 2008). Acest lucru face ca o sarcină solicitantă pentru țintă sau selecție, împreună cu dificultatea expresiei funcționale a proteinelor transmembranare, cum ar fi ORs, pentru a afișa un număr mare de liganzi. O aproximare a proprietăților importante în ambele OBP și rup sunt rezumate în tabelul 3.
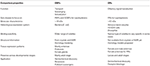
Tabelul 3., Comparație aproximativă a ORs insecte și OBPs în funcție de proprietăți.
Alte Perspective
funcționale caracterizarea de insecte ORs, precum și dovedit roluri în insecte olfactive au vărsat luminile de pe sensibilitatea și specificitatea acestor insecte-proteine specifice. Aceste progrese vor spori și mai mult fezabilitatea lor ca obiective de combatere a dăunătorilor prin înțelegerea mecanismelor de recunoaștere moleculară și a interacțiunilor combinatorii cu OBPs., Pe de altă parte, actualul efort masiv în identificarea și obligatoriu caracterizarea OBPs în mai multe agricol important de specii de insecte va continua și de a oferi mai multe informații despre funcțiile lor în fiziologia insectelor. Astfel, această revizuire propune ca principal avantaj pentru OBPs peste ORs, disponibilitatea de cristal 3D și RMN structuri, care cu în aval abordări, cum ar fi omologie de modelare (când este necesar), molecular docking și dinamică moleculară, ar rafina căutarea bioactive produse chimice., Acest ultim în completare cu ligand de afinitate de măsurare va accelera studiu de insecte OBPs să fie reconsiderate ca obiective pentru semiochemical descoperirea și instrumente pentru design super-liganzi în combaterea dăunătorilor de management.apariția și dezvoltarea rezistenței insecticidelor la insectele dăunătoare au condus la cercetarea intensivă a olfacției insectelor și a mecanismelor implicate în procesarea neuronală. Este bine stabilit că un număr de receptori și enzime din SNC de insecte sunt țintele pentru dezvoltarea rezistenței la insecticide (Figura 1)., S-a demonstrat că acetilcolinesteraza (AChE) în formă solubilă asigură rezistența la insecticidele organo-fosfor și carbamat, acționând ca bioscavengeri (Lee et al., 2015). În mod similar, mai multe insecticid mecanisme de rezistență au fost demonstrate în afide M. persicae, care implică carboxylesterases, canalele de sodiu, acidul γ-aminobutiric (GABA) și nAChR (Bass et al., 2014)., Ca și componente importante în insecte periferia sistemului nervos și actori-cheie în insecte comportamente, atât de insecte OBPs și Rup reprezintă o alternativă obiective pentru identificarea compușilor cu semiochemical activitate (sau efect agonist) și instrumente pentru design puternic antagoniști pentru a spori dorit răspunsurile comportamentale de insecte dăunătoare și de a reduce utilizarea de insecticide și rezistență ulterioară.
contribuțiile autorului
HV a scris secțiuni despre structura OBP și rup, tabele dezvoltate și figura., J-JZ a conceput ideea pentru articolul de revizuire, a scris secțiunea principală, cum ar fi introducerea, funcția și structura OBPs, precum și compararea OR-OBP.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
mulțumiri
autorii doresc să mulțumească FONDECYT 3170433., J-JZ este recunoscător pentru sprijinul financiar din partea Universității normale de Nord-Est și a Universității Jilin, China pentru concediul său sabatic de a studia în China. Rothamsted Research primește un grant cu finanțare suplimentară de la biotehnologie și științe Biologice (BBSRC), Marea Britanie.
Gonzalez, F., Witzgall, P., and Walker, W. B. (2017). Antenare transcriptomes de trei tortricid molii dezvăluie prezumtiv conservate chemosensory receptorii sociale și habitat olfactiv indicii. Sci. Rep. 7, 1-12. doi: 10.,1038/srep41829
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Kaissling, K. E. (2013). Cinetica răspunsurilor olfactive ar putea depinde în mare măsură de interacțiunea odorant-receptor și de dezactivarea odorantului postulată pentru detectoarele de flux. J. Comp. Physiol. Neuroetol. Se Comportă Neural Senzorial. Physiol. 199, 879–196. doi: 10.1007/s00359-013-0812-z
PubMed Abstract | CrossRef Textul Complet | Google Scholar