Proiectarea și construcția de studiu randomizat biblioteci pentru testari Cas9 PAM preferințele
PAM biblioteci care conțin randomizat secvente de ADN imediat în aval de o secvență de ADN complementar cu distanțier de un ghid de ARN au fost generate și utilizate pentru a determina empiric PAM recunoaștere de Tip II Cas9 endonucleaza (Fig. 1)., Odată cu fixarea secvenței țintă a distanțierului ARN de ghidare, bazele randomizate servesc ca substrat pentru citirea directă a specificității Pam a Endonucleazei Cas9. Secvențele randomizate au fost introduse într-un vector ADN plasmidic în regiunea PAM a unei secvențe țintă protospacer care demonstrează omologia perfectă a distanțierului ARN T1 (CGCUAAAGAGGAAGAGGACA). Au fost generate două biblioteci care cresc în dimensiune și complexitate de la cinci perechi de baze randomizate (1,024 combinații potențiale PAM) la șapte perechi de baze randomizate (16,384 combinații potențiale PAM)., Randomizarea bibliotecii 5 bp a fost introdusă prin sinteza unei singure oligonucleotide care conține cinci reziduuri aleatorii. Oligonucleotida monocatenară a fost transformată într-un șablon dublu catenar prin PCR (fișierul suplimentar 1: Figura S1A), clonată în vectorul plasmidic (fișierul suplimentar 1: Figura S1B) și transformată în E. coli, așa cum este descris în secțiunea Metode., Pentru a asigura randomizarea optimă în biblioteca PAM de 7 bp, dimensiunea și complexitatea bibliotecii au fost reduse prin sintetizarea a patru oligonucleotide, fiecare conținând șase reziduuri aleatorii, plus un al șaptelea reziduu fix care conține G, C, A sau T, respectiv. Fiecare dintre cele patru oligonucleotide au fost transformate separat în ADN dublu catenar, clonate în vector pTZ57R / T așa cum este descris în secțiunea Metode și transformate în E. coli așa cum este descris pentru biblioteca 5 bp., După transformare, ADN-ul plasmidic a fost recuperat și combinat din fiecare dintre cele patru biblioteci PAM de 6 bp pentru a genera o bibliotecă Pam de 7 bp randomizată care cuprinde 16,384 de combinații posibile Pam. Pentru ambele biblioteci, încorporarea de dezordine a fost validată de adânc secvențiere; examinarea nucleotide compoziție la fiecare poziție a PAM regiune folosind o poziție de frecvență matrix (MFP) (secțiunea Metode și ) (fișier Suplimentar 1: Figura S2A și B)., Distribuția și frecvența fiecărei secvențe PAM în 5 bp și 7 BP randomizat biblioteca Pam sunt prezentate în fișier suplimentar 1: cifrele S3 și S4, respectiv.
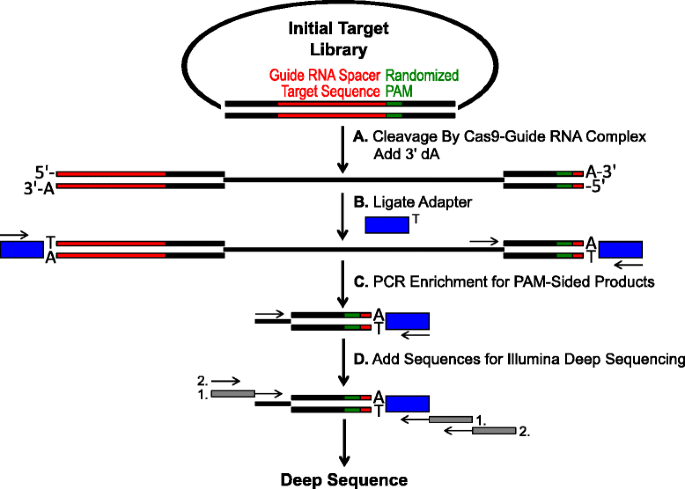
Schema de identificare pentru PAM preferințele de Cas9 decolteu in vitro. o bibliotecă plasmidă inițială cu Pam randomizat (cutie verde) este scindată cu complexul Cas9 și se adaugă 3 ” console dA. adaptoarele B cu consola 3 ‘ dT (cutie albastră) sunt ligate la ambele capete ale produsului de clivaj., c primeri sunt utilizate pentru a îmbogăți pentru PAM-verso clivate produse prin PCR., d După PCR îmbogățire, fragmente de ADN sunt purificate și Illuminati compatibil ancore și codurile de bare sunt ‘coada-pe’ prin intermediul a două runde de PCR (cutii gri) și Illuminati adânc ordonate
Testari Cas9 PAM preferințele
randomizat PAM biblioteci descrise în secțiunea anterioară au fost supuse digestie in vitro cu diferite concentrații de recombinare Cas9 proteine preîncărcat cu ghid ARN, în scopul de a analiza Cas9 endonucleaze PAM preferințele într-un mod dependent de doză., După digestie cu Cas9-ghid de ARN ribonucleoprotein (RNP) complexe, PAM secvență combinații de studiu randomizat PAM bibliotecă, care a sprijinit clivaj au fost capturați de către ligatura adaptoare pentru gratuit-capetele de plasmidă molecule de ADN monocatenar de Cas9-ghid de ARN complexe (Fig. 1a și b). Pentru a promova eficient ligatura și captura din capete despicat, bont-a încheiat ADN dublu catenar taie generate de Cas9 endonucleaza a fost modificat pentru a conține un 3′ dA consola si adaptoare fost modificat pentru a conține un complementare 3′ dT consola., Pentru a genera cantități suficiente de ADN pentru secvențiere, fragmentele de ADN care adăpostesc secvența PAM care susțin clivajul au fost amplificate PCR folosind un primer în adaptor și un altul direct adiacent regiunii PAM (Fig. 1C). Bibliotecile PCR amplificate Cas9 Pam rezultate au fost transformate în șabloane ampli-seq (Fig. 1D) și o singură citire profundă secvențiate din partea adaptorului amplicon., Pentru a asigura o acoperire adecvată, la Cas9 PAM biblioteci au fost esalonate la o adâncime de cel puțin cinci ori mai mare decât diversitatea în primele randomizat PAM library (5,120 și 81,920 citește pentru 5 și 7 bp PAM randomizat biblioteci, respectiv). PAM secvențe au fost identificate din care rezultă secvența de date de doar selectarea celor citește conțin o perfectă 12 nt secvență meci flancat de fiecare parte a 5 sau 7 nt PAM secvență (în funcție randomizate PAM bibliotecă utilizate); captarea numai cei PAM secvențe care rezultă din perfect Cas9-ghid ARN-ul țintă de recunoaștere și decolteu., Pentru a compensa părtinirea inerentă în bibliotecile Pam randomizate inițiale, frecvența fiecărei secvențe PAM a fost normalizată la frecvența sa în biblioteca de pornire. Deoarece testul descris aici captează direct secvențele Pam scindabile Cas9, modelarea probabilistică a fost utilizată pentru a calcula consensul PAM pentru fiecare proteină Cas9. Acest lucru a fost realizat prin evaluarea probabilității de a găsi fiecare nucleotidă (G, C, A sau T) la fiecare poziție a secvenței PAM independent folosind o matrice de frecvență de poziție (PFM) (secțiunea Metode și )., Probabilitățile rezultate au fost apoi vizualizate ca un WebLogo .pentru a examina înclinația pentru fals pozitive în test, adăugarea complexelor Cas9 RNP în etapa de digestie a fost omisă (Fig. 1A) și testul a fost efectuat prin etapa de îmbogățire PCR (Fig. 1C). După cum se arată în fișierul suplimentar 1: Figura S5A, nu au fost detectate produse de amplificare în absența complexelor ARN de ghidare Cas9. Astfel, indicând faptul că incidența fals pozitive este scăzută și nu contribuie în mod semnificativ la rezultatele testului.,
PAM preferințele de Streptococcus pyogenes și Streptococcus thermophilus (CRISPR3 și CRISPR1 sisteme) Cas9 proteine
În scopul de a valida testul PAM preferințele de Streptococcus pyogenes (Spy) și Streptococcus thermophilus CRISPR3 (Sth3) Cas9 proteine, a căror PAM secvență cerința au fost raportate anterior , au fost examinate. In vitro digera au fost efectuate cu 1 µg (5,6 nM) 5 bp randomizat PAM biblioteca de la două concentrații de 0,5 și 50 nM, de pre-asamblate Spion sau Sth3 Cas9 proteine, crRNA, și tracrRNA ROMSILVA complexe pentru 1 h în 100 µL de reacție volum., În funcție de frecvența lor în 5 bp randomizat PAM bibliotecă, Spion și Sth3 Cas9 PAM secvențe (NGG și NGGNG, respectiv) au fost la concentrațiile finale de 0,40 nM și de 0,11 nM în digestie, respectiv. Membrii Bibliotecii Pam randomizate care conțineau secvențe PAM care susțineau scindarea au fost capturați și identificați așa cum este descris în secțiunea anterioară. Ca un control negativ, biblioteca Pam randomizată neavizată inițială a fost supusă secvențierii și analizei PFM alături de acele biblioteci expuse complexelor RNP Cas9., Așa cum se arată în fișier Suplimentar 1: Figura S5B și C, nici o secvență de preferințele exista în absența Cas9 ROMSILVA complex digestia ca evident de o apropiere de distribuție perfectă de fiecare nucleotide la fiecare poziție a PAM în MFP masă și lipsa de conținut informativ în WebLogo pentru control. Acest lucru este în constrast stark cu Fig. 2a și b care ilustrează compoziția secvențelor derivate din biblioteci digerate cu complexe RNP Spy și Sth3 Cas9. Examinarea paginilor derivate din MFP (Fig ., 2a și b) dezvăluie, de asemenea , prezența preferințelor canonice Pam pentru proteinele Spy și Sth3 Cas9, NGG și NGGNG, respectiv. Deși PAM preferințele raportat pentru Spionaj și Sth3 Cas9 proteinele sunt observate în ambele 0,5 nM 50 nM digera, este, în general, extinderea în specificitate sub 50 nM digera condiții. Acest lucru este cel mai evident în poziția 2 pentru proteina Spy Cas9 unde frecvența unui reziduu non-canonic crește dramatic (Fig. 2a)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
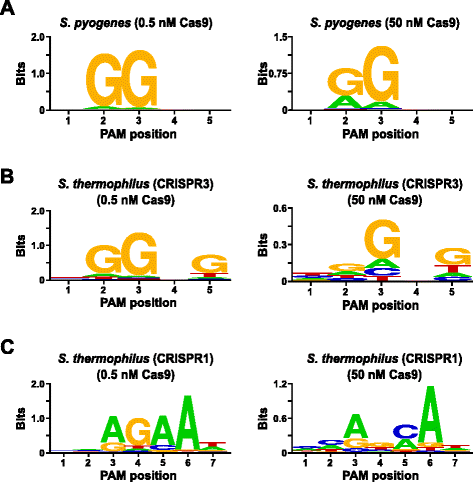
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., Frecvența de nucleotide la fiecare PAM poziția de independent a fost calculată folosind-o poziție de frecvență matrix (MFP) și reprezentate grafic ca o WebLogo
în Continuare de validare a testului a fost realizat prin examinarea PAM preferințele pentru Streptococcus thermophilus CRISPR1 (Sth1) Cas9 proteine a căror PAM specificitatea a fost raportat de a extinde de la 7 bp . Folosind 1 µg (5,6 nM) de 7 bp randomizat PAM biblioteca ca șablon, Sth1 Cas9-ghid de ARN digestie au fost efectuate la două concentrații de 0,5 nM 50 nM, de RNP complex cum este descris mai sus., Ca controale, complexele RNP Spy și Sth3 Cas9 au fost, de asemenea, utilizate pentru a digera biblioteca PAM randomizată 7 bp, dar numai la o singură concentrație complexă RNP de 0,5 nM. În funcție de frecvența în 7 bp randomizat PAM bibliotecă, PAM secvențe raportate anterior pentru Sth1 (NNAGAAW), Spion (NGG), și Sth3 (NGGNG) au fost la final concentrații de 0,01 nM, 0.22 nM, și 0,05 nM, respectiv., Așa cum se arată în fișier Suplimentar 1: Figura S6A și B, PAM preferințele pentru Spionaj și Sth3 Cas9 proteine generate folosind 7 bp bibliotecă au fost aproape identice cu cele produse cu 5 bp biblioteca furnizarea de dovezi puternice pentru reproductibilitatea testului. Preferințele PAM pentru proteina Cas9 Sth1 s-au potrivit, de asemenea , cu cea raportată anterior, NNAGAAW, la concentrația complexului ARN de ghidare Cas9-0.5 nM (Fig. 2c)., Similare pentru a Spiona și Sth3 Cas9 proteine, Sth1 Cas9 a fost capabil de a smulge un set mai divers de PAM secvențe în reacții care conțin o concentrație mai mare de Cas9-ARN ghid complex (50 nM), cea mai izbitoare a fost marcat de pierdere a G reziduu cerința de la poziția 4 și aproape egale, preferința pentru o C și tensiunea la poziția 5 (Fig. 2c). Acest lucru a dus la un consens PAM diferit de cel obținut la concentrații mai mici.,
Pentru a examina dacă PAM specificitatea este independent de tipul de ARN ghid, duplexed crRNA:tracrRNA sau sgRNA , Spion, Sth3, și Sth1 Cas9 PAM preferințele au fost examinate, de asemenea, folosind un binar Cas9 și sgRNA ROMSILVA complexe. Digestia a fost efectuată la o singură concentrație complexă RNP de 0,5 nM și analiza preferințelor PAM a fost efectuată așa cum este descris mai sus. Așa cum se arată în fișier Suplimentar 1: Figura S7A, B, și C, PAM preferințele au fost aproape identice indiferent de tipul de ghid ARN folosit; fie o crRNA:tracrRNA duplex sau sgRNA., În plus, pentru a confirma că PAM specificitatea nu este foarte mult influențată de compoziția ADN-ului țintă sau spacer secventa, secventa pe partea opusă a 5 sau 7 bp randomizat biblioteca a fost vizat pentru decolteul cu un alt distanțier; T2-5 (UCUAGAUAGAUUACGAAUUC) pentru 5 bp bibliotecă sau T2-7 (CCGGCGACGUUGGGUCAACU) pentru 7 bp bibliotecă. Spion și Sth3 Cas9 proteine preîncărcat cu sgRNAs vizând secvența T2 au fost folosite pentru a interoga 5 bp randomizat PAM bibliotecă în timp ce Sth1 Cas9-T2 sgRNA complexe au fost utilizate pentru a digera 7 bp randomizat PAM bibliotecă., Preferințele PAM au fost evaluate așa cum este descris mai sus. Preferințele PAM pentru toate cele 3 proteine Cas9 au fost aproape identice, indiferent de secvența ADN-ului distanțier și țintă (fișierul suplimentar 1: Figura S8A, B și C).
Identificarea sgRNA și PAM preferințele pentru Brevibacillus laterosporus Cas9 proteine
Să empiric examina PAM preferințele pentru o Cas9 proteine a căror PAM a fost nedefinit, o uncharacterized Tip II-C CRISPR-Cas locus de Brevibacillus laterosporus tulpina SSP360D4 (Blat) a fost identificat prin căutarea internă DuPont Pioneer baze de date pentru Cas9 orthologues., Locus (aproximativ 4.5 kb) conținea o cas9 gene capabile de codare o 1,092 polipeptidice, un CRISPR matrice care cuprinde șapte repeta-distanțier de unități doar în aval de cas9 gene și o tracrRNA codare regiune situată în amonte de cas9 gene cu omologie parțială la CRISPR matrice se repetă (Fig. 3a). Lungimea repetării și a distanțierului (36 și 30 bp, în consecință) este similară cu alte sisteme CRISPR-Cas de tip II, cu cinci din cele opt repetări care conțin 1 sau 2 mutații bp (Fig. 3B și fișier suplimentar 1: Figura S9)., Alte gene găsite de obicei într-un locus CRISPR-Cas de tip II au fost fie trunchiate (cas1), fie lipsă (Fig. 3a).
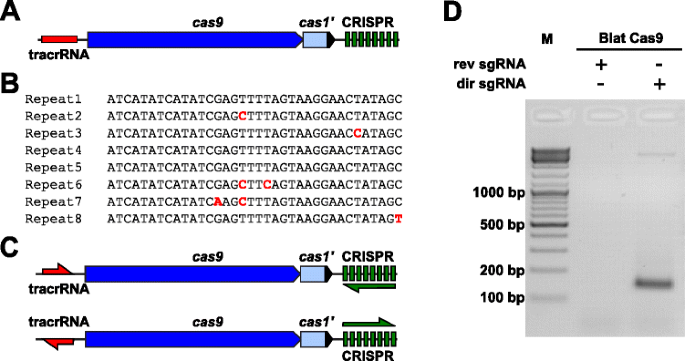
Identificare de Tip II CRISPR-Cas elemente în Brevibacillus laterosporus SSP360D4 CRISPR-Cas sistem. a o ilustrare a regiunii ADN genomic din sistemul CRISPR-Cas de tip II de la Brevibacillus laterosporus SSP360D4. b Comparație de Tip II CRISPR matrice repetați secvențele identificate în Brevibacillus laterosporus SSP360D4., c „directe” și „invers” tracrRNA și CRISPR matrice transcripțional scenarii de Tip II CRISPR-Cas sistem de Brevibacillus laterosporus SSP360D4. d-Un gel de agaroză cu produse de reacție, indicând faptul că numai „directe” sgRNA (dir sgRNA), dar nu „invers” sgRNA (rev sgRNA) suport plasmidă biblioteca clivaj în combinație cu Cas9 endonucleaze originare din Brevibacillus laterosporus SSP360D4
ghidul ARN cerință pentru Blat Cas9 proteine a fost determinată prin generarea de două sgRNA variante., Aceste variante au fost generate pentru a ține cont atât de posibilele scenarii de Expresie sens sau anti-sens ale matricei tracrRNA, cât și de CRISPR (Fig. 3c) și folosit pentru a sonda ce scenariu de expresie a sprijinit activitatea de clivaj a Blat Cas9 în biblioteca Pam randomizată. Un singur ghid RNAs au fost concepute mai întâi prin identificarea limitelor de prezumtiv tracrRNA molecule de analiza regiuni care au fost parțial complementare 22 nt 5′ terminal al repetare (anti-repetare)., Apoi, pentru a determina capătul 3 ‘ al tracrRNA, s-au folosit posibile structuri secundare și terminatoare pentru a prezice regiunea de terminare în fragmentul din aval. Acest lucru a fost realizat prin screening-ul pentru prezența secvențelor de terminare Rho independente în ADN-ul care înconjoară anti-repetarea similară cu cea descrisă în Karvelis et al. , transformarea ADN-ului înconjurător în secvența ARN și examinarea structurilor rezultate folosind UNAfold ., Rezultanta sgRNAs au fost concepute pentru a conține un T7 polimeraza transcriere inițierea de recunoaștere a semnalului de la 5′ end urmat de 20 nt țintă secvența de recunoaștere, 16 nt de crRNA repet, 4 nt auto-pliere ac de păr buclă, și anti-repetați secvența complementară regiune repeta de crRNA urmată de celelalte 3′ parte din prezumtiv tracrRNA. Varianta sgRNA care conține o presupusă tracrna transcrisă în aceeași direcție cu gena cas9 (Fig. 3c) este numit „directe” sgRNA, în timp ce sgRNA conține tracrRNA transcrise în direcția opusă un „invers” sgRNA., Cincizeci nM de Blat Cas9 sgrna RNP complex, pre-încărcate fie cu sgrnas „directe” sau „inversă”, respectiv, au fost incubate cu 1 µg (5.6 nM) din 7 bp randomizat biblioteca PAM. După digestia bibliotecii și adăugarea consolelor 3’ dA, adaptoarele au fost ligate și produsele de clivaj au fost amplificate PCR (Fig. 1). Analiza produșilor de reacție prin electroforeză pe gel de agaroză a arătat că „directe” sgRNA, dar nu „invers” sgRNA sprijinit plasmidă bibliotecă de clivaj (Fig. 3d). Secvența și structura secundară prezisă a sgRNA „directă” sunt prezentate în fișierul suplimentar 1: Figura S10.,
După stabilirea ARN ghid pentru Blat Cas9, PAM identificarea s-a realizat în mod similar cu cea descrisă mai sus pentru Spion, Sth3, și Sth1 Cas9 proteine împotriva 7 bp randomizat PAM bibliotecă cu două concentrații de 0,5 și 50 nM, de pre-asamblate Blat Cas9 „directe” sgRNA ROMSILVA complexe. Așa cum se arată în Fig. 4a, MFP WebLogo PAM consens pentru Blat Cas9 proteine sub 0.5 nM digera condiții fost NNNNCND (N = G, C, a, sau T; D = a, G sau T) cu o preferință puternică pentru un C la poziția 5 din PAM secvență., O preferință moderată pentru un A a fost observată în poziția 7 și ușoare preferințe pentru a C sau T în poziția 4 și G, C sau A peste T în poziția 6 au fost, de asemenea, observate la examinarea atentă a tabelului MFP (fișier suplimentar 1: Figura S11). În mod similar, pentru a Spiona, Sth3, și Sth1 Cas9 proteine, PAM specificitatea lărgește ca Cas9-sgRNA complex de concentrare crește. Acest lucru este cel mai evident în poziția 5, unde o proporție mai mare de secvențe PAM care conțin o scindare a suportului de reziduuri A la 50 nM comparativ cu condițiile de digerare de 0,5 nM.
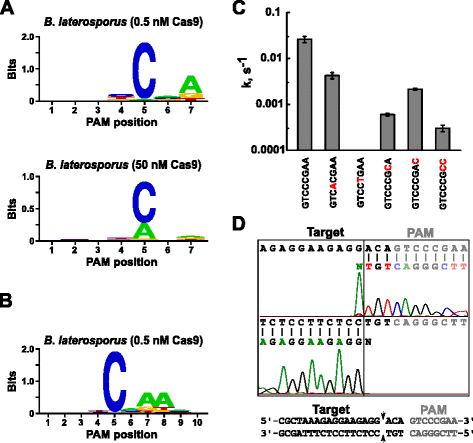
PAM preferințele și decolteul poziții de Brevibacillus laterosporus SSP360D4 (Blat) Cas9 enzime. Blat Cas9 PAM preferințe atunci când 1 µg de biblioteca ADN-ul a fost clivat cu 0,5 nM sau 50 nM Cas9-sgRNA complex (o), extins la poziția 10 prin schimbarea protospacer țintă de 3 bp (b). Frecvența nucleotidelor la fiecare poziție PAM a fost calculată independent folosind o matrice de frecvență de poziție (PFM) și reprezentată grafic ca WebLogo ., c ratele de scindare a substraturilor de ADN plasmidic supracoilat care conțin mutații (prezentate în roșu) în secvența PAM GTCCCGAA. Toate punctele de date sunt valori medii din ≥3 experimente independente. Barele de eroare sunt date ca S. D. d Run-off succesiunea de la ambele simț și anti-sens direcții de plasmide de ADN monocatenar, cu Blat Cas9
Din Blat Cas9 poate accepta orice de bază în primele trei poziții ale sale PAM secvență (Fig. 4A), distanțierul T1 a fost deplasat cu trei nucleotide în direcția 5′ pentru a permite extinderea identificării PAM de la 7 la 10 bp., A schimbat T1 distanțier, T1-3 (AAACGCUAAAGAGGAAGAGG), a fost încorporat în Blat „directe” sgRNA și PAM identificarea s-a realizat după cum este descris anterior pentru Spionaj, Sth3, Sth1, și Blat Cas9 proteine. Analiza preferințelor PAM a relevat specificitatea PAM pentru blat Cas9 poate fi extinsă la poziția 8 unde există o preferință moderată pentru un A suplimentar (Fig. 4b).
specificitatea PAM pentru blat Cas9 a fost confirmată prin generarea de plasmide care să conțină mutații în cele mai conservate reziduuri ale PAM (Fig. 4c)., Înlocuirea nucleotidei C la poziția 5 a eliminat scindarea ADN-ului plasmidic confirmând rolul său cheie în recunoașterea Blat Cas9 Pam. Înlocuirea unei nucleotide la pozițiile 7 și 8 a redus semnificativ (43× și respectiv 12×) rata de scindare a plasmidei supracoilate, indicând, de asemenea, importanța acestor nucleotide în recunoașterea Pam Cas9 Blat.,
Pentru a identifica ADN-ul țintă clivaj poziții pentru Blat Cas9 proteine, o plasmidă care conțin 20 bp regiunea de potrivire distanțier T1 urmat de un PAM secvență, GTCCCGAA, care se încadrează în PAM consens pentru Blat Cas9, NNNNCNDD, a fost generată și digerate cu Blat Cas9-ghid de ARN ribonucleoprotein complexe. Secvențierea directă a ADN-ului a fost utilizată pentru a determina capetele moleculei ADN liniare generate de Complexul RNP Blat Cas9. Rezultatele secvenței au confirmat că scindarea ADN-ului plasmidic a avut loc în protospacerul 3 nt 5′ al secvenței PAM (Fig., 4D) similar cu cel observat pentru proteinele Spy, Sth3 și Sth1 Cas9 .
În planta genomului editare folosind Blat Cas9 și sgRNA
în Urma elucidarea sgRNA și PAM preferințele pentru Blat Cas9, porumb optimizat Cas9 și sgRNA expresie casete au fost generate în planta de testare cum a fost descris anterior pentru S. pyogenes cas9 gene și sgRNA . Pe scurt, gena Blat cas9 a fost codonul de porumb optimizat și intronul 2 al genei St-LSI de cartofi a fost introdus pentru a perturba expresia în E. coli și a facilita îmbinarea optimă în planta (fișier suplimentar 1: Figura S12)., Localizare nucleară de Blat Cas9 proteine din porumb celule a fost facilitat prin adăugarea de ambele amino și carboxil-terminal nucleare locații semnale, SV40 (MAPKKKRKV) și Agrobacterium tumefaciens VirD2 (KRPRDRHDGELGGRKRAR), respectiv (fișier Suplimentar 1: Figura S12). Gena Blat cas9 a fost exprimată constitutiv în celulele vegetale prin legarea cas9 optimizată la un promotor Ubiquitin de porumb și la terminatorul pinII într-un vector ADN plasmidic., Pentru a conferi eficienta sgRNA exprimare în porumb celule, un porumb U6 polimeraza III promotor și terminator (TTTTTTTT) au fost izolate și s-a lipit de 5′ și 3′ capetele de modificat Blat sgRNA codificare secvența ADN, respectiv (fișier Suplimentar 1: Figura S13). Modificat Blat sgRNA conținea două modificări de cea utilizată în studiile in vitro; o T a G modificări la poziția 99 și a T a C modificare la poziția 157 din sgRNA (fișier Suplimentar 1: Figura S13). Modificările au fost introduse pentru a elimina potențialele semnale premature de terminare a polimerazei U6 III în Blat Sgrna., Modificările introduse pentru a avea un impact minim asupra structurii secundare a sgRNA comparativ cu versiunea utilizată în studiile in vitro (date care nu sunt prezentate).
Pentru a compara cu precizie mutatii de eficiență care rezultă din imperfectă non-omoloage end-unirea (NHEJ) de reparare a ADN-ului dublu-strand (DSBs) care rezultă din Spy si Blat Cas9 clivaj, protospacer identice genomice țintă site-uri au fost selectate prin identificarea obiectivelor cu Spy si Blat Cas9 compatibil PAMs, NGGNCNDD., Secvențele distanțiere identice au fost selectate pentru Blat și Spy Cas9 prin captarea secvenței 18 la 21 nt imediat în amonte de PAM. Pentru a asigura o expresie optimă a polimerazei U6 III și a nu introduce o nepotrivire în distanțierul sgRNA, toate secvențele țintă au fost selectate pentru a se termina în mod natural într-un G la capătul lor 5′. Obiectivele au fost identificate și selectate în exonul 1 și 4 din porumb fertilitate gene Ms45 și într-o regiune în amonte de porumb liguleless-1 gene.,activitatea mutațională a Blat Cas9 la porumb a fost examinată prin transformarea biolistică a embrionilor imaturi de porumb vechi de 10 zile (ime) cu vectori ADN care conțin gene cas9 și sgrna. Blat și Spy echivalent Cas9 și vectorii de Expresie sgRNA au fost introduse în mod independent, în ime de porumb Hi-tip II prin transformarea pistolului de particule similar cu cel descris în . Deoarece transformarea pistolului de particule poate fi foarte variabilă, o casetă de expresie a ADN-ului marker vizual, Ds-Red, a fost, de asemenea, co-livrată cu vectorii de Expresie Cas9 și Sgrna pentru a ajuta la selectarea ime uniform transformate., În total, trei replici de transformare au fost efectuate pe 60-90 IME și 20-30 dintre ime cele mai uniform transformate din fiecare replicat au fost recoltate la 3 zile după transformare. ADN-ul genomic total a fost extras și regiunea din jurul site-ului țintă a fost amplificată prin PCR și amplicons secvențiat la o adâncime de citire mai mare de 300.000. Rezultate citi au fost examinate pentru prezența mutațiilor la temperatura site-ul de clivaj prin comparație cu experimente de control unde sgRNA ADN expresia caseta a fost omis din transformare. Așa cum se arată în Fig., 5A, mutațiile au fost observate la locul așteptat de clivaj pentru Blat Cas9, cele mai răspândite tipuri de mutații fiind inserțiile sau ștergerile unei singure perechi de bază. Modele similare de reparare au fost observate și pentru proteina Spy Cas9 (fișier suplimentar 1: Figura S14 și ). Activitatea mutațională pentru Blat Cas9 a fost robustă la două dintre cele trei site-uri testate și a depășit-o pe cea a spionului Cas9 la site-ul țintă Ms45 exon 4 cu aproximativ 30 % (Fig. 5b).
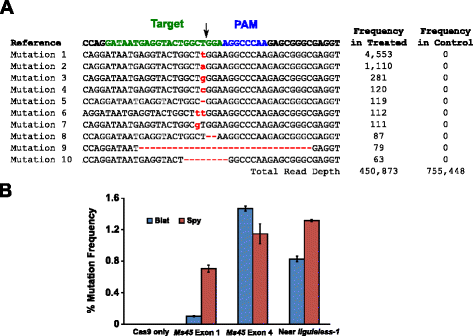
Brevibacillus laterosporus Cas9 promovează NHEJ mutații în porumb. un Top 10 cele mai răspândite tipuri de mutații NHEJ detectate cu Blat Cas9 în exonul 4 al genei Ms45. O săgeată neagră indică locul așteptat de clivaj; mutațiile sunt evidențiate cu roșu; fontul cu litere mici indică o inserție; ” – ” indică o ștergere. B Compararea frecvențelor de mutație Spy și Blat Cas9 Nhej la trei site-uri țintă identice protospacer în porumb. Mutațiile NHEJ au fost detectate prin secvențiere profundă la 3 zile după transformare., Barele de eroare reprezintă sem, n = 3 transformări ale pistolului de particule. Numai Cas9 este controlul negativ și reprezintă media (în toate cele trei site-uri țintă) frecvența de fond a mutațiilor rezultate din amplificarea și secvențierea PCR