Även om flyktiga föreningar med agonist aktivitet har visats mot de yttersta Randområdena, i ett specifikt ämne med stark agonist effekt på myggor, 2-(4-Etyl-5-(pyridin-3-yl)-4H-1,2,4-triazol-3-ylthio)-N-(4-ethylphenyl)acetamid (VUAA1), har öppnat forskning (Jones et al., 2011; Taylor et al., 2012). Senare studier av Taylor et al., (2012) styrkt VUAA1-och oljebaserade kemikalier, såsom VUAA4, kunna öka sin agonist effekten av 10-vik på ORco från A. gambiae, H. virescens, och Harpegnathos saltator. Intressant rapporterade författarna att någon förändring av amid-substituenter kommer att orsaka en fullständig förlust av agonistisk aktivitet. Detta ger användbara insikter om agonisternas strukturella krav och strukturaktivitetsförhållandet mellan VUAA-analoger och ORs. Slutligen, trots den förbättrade agonistiska aktiviteten hos VUAA chemicals, dess relativt höga molekylvikt (367,47 g mol−1 för VUAA1) vs., flyktiga agonister, såsom bombykol (238.42 g mol-1), Gör en direkt Flyktig leverans av VUAA något inte möjligt. Med detta i åtanke representerar sökandet efter mindre strukturella analoger ett intressant forskningsfokus.
Antagonism på ORco-subenheten
tillsammans med studien av VUAA-relaterade analoger som kan fungera som starka agonister har blockeringen av ORco av antagonister också uppstått för att styra semiokemikalier och bekämpningsmedel., Alltså, en strukturell analog av VUAA1, VU0183254 (2-(4-Etyl-5-furan-2-yl-4H-triazol-3-ylsulfanyl)-1-fenotiazin-10-yl-ethanone), rapporterades att hämma ORco svar, i egenskap av allosteric modulator i A. gambiae och störa erkännande av agonister som eugenol av komplexa OR65/ORco (Jones et al., 2012). Andra VUAA-strukturella analoger har också rapporterats som antagonister. Ett exempel är N-,2-substituerade triazolothioacetamide föreningar OLC3 och OLC12 som stör ORco svar på ett liknande sätt i C. quinquefasciatus, A. gambiae, D. melanogaster och O., nubilalis, vilket tyder på en bevarad bindningsplats i ORco (Chen och Luetje, 2012). Med tanke på hämningen av ORco som en lovande strategi för att störa beteenden hos insekter verkar det som att efterföljande ansträngningar bör inriktas på föreningarna med lägre molekylvikt än VUAA-härledda antagonister. Till exempel rapporterades OX1a (232 g mol−1), tryptamin (160.22 g mol−1) och isopropylcinnamat (190.24 g mol−1) ha antagonistisk effekt på ORco (Chen och Luetje, 2013, 2014; Tsitoura et al., 2015) med ungefär halv eller mindre molekylvikt än VUAA1., Ändå bör framtida användning av dessa antagonister studeras noggrant, eftersom blockeringen av den konserverade ORco kan påverka inte bara skadliga insekter utan också fördelaktiga.
förutom den antagonistiska effekten som undersöktes in vitro stöder bevisen på beteendenivå tanken att strukturella analoger av feromoner kan fungera som antagonister. Till exempel, Sellanes et al., (2010) rapporterade hämningen av sexuellt svar i honeydew moth Cryptoblabes gnidiella när de strukturella analogerna, (Z)-9-tetradecenylformiat och (Z)-11-hexadecenylformiat, tillsattes till syntetiskt kön feromon, (Z)-11-hexadecenal och (Z)-13-oktadecenal, i vindtunneltester. Denna feromonantagonist effekt bekräftades senare i fältanalyser, där fångsten av C. gnidiella-män minskade i ett dosberoende mönster. Feromonantagonismen har också rapporterats för B. mandarina, en förfader till B. mori (Daimon et al., 2012)., Deras fynd bekräftar bombykol som kön feromon och bombykal och bombykylacetat som antagonister, vilket starkt hämmade attraktionen hos män i fält till kön feromon bombykol. Mer nyligen rapporterades bevis på feromonantagonism för snout moth Herpetogramma submarginale. När (Z)-13-hexadecenol tillsattes till sitt kön feromon, (Z)-13-hexadecenylacetat, minskade signifikant antalet män som fångats i fält (Yan et al., 2015)., Feromonantagonismen verkar baserat på skillnaderna i kemisk funktionell grupp som alkoholer, aldehyder och estrar beroende på insektsarterna. Den antagonistiska effekten av dessa strukturella analoger kan dock inte bero på ORco-hämning utan på ORX: s specificitet mot antagonister. En ny studie tyder på att Or16 av Helicoverpa armigera specifikt kan känna igen feromonantagonisten, (Z)-11-hexadecenol (Chang H. et al., 2017). Författarna stödde OR16S specifika roll med tanke på att H., armigera Honor avger antagonistföreningen tillsammans med sin sexferomon ((Z)-11-hexadecenal och (Z)-9-hexadecenal) som en strategi för att undvika icke-optimal parning med omogna män. Enastående, när OR16 slogs ner av genomredigeringstekniken CRISPR/Cas9 och H. armigera-män testades genom elektrofysiologi och beteendeanalyser, registrerades inget EAG-svar och män försökte para sig med omogna kvinnor.
luktreceptorer vs., Bindningsproteiner: fördelar och nackdelar för Insektspesthantering
När det gäller OBPs representerar ligandspecificiteten och mekanismerna för OBPs kontroversiella aspekter, vilket verkar starkt beroende av de metoder som används för mätning av ligand affinitet. Det har till exempel rapporterats att PBPs, såsom de från nattfjärilar’ S. xylostella och Eogystia hippophaecolus, kan binda både sex feromon komponenter och analoger (Sun et al., 2013a; Hu m.fl., 2018). Detta tyder på att nedströms spelare som ORs kan förbättra specificiteten och känsligheten hos luktmottagning., Nya bevis stöder att samuttryck av PBPs och PRs kan öka känsligheten mot feromoner. Till exempel användes flera kombinationer från PR1-4 och PBP1-4 för att testa deras svar på sexferomonkomponenterna i moth Chilo suppressalis (Chang et al., 2015). Författarna fann en signifikant ökning av känsligheten för svar mot (Z)-11-hexadecenal när PR4 och PR6 uttrycktes tillsammans med PBP4., Även om interaktionen mellan dessa proteiner kan uppstå en ny nivå av forskning som skadedjursbekämpningsmål, belyser den olika parningen av PRs och PBPs komplexiteten hos det olfaktoriska systemet i insekter, vilket gör tillvägagångssättet en svår uppgift för en stor uppsättning föreningar och proteiner att testa. Trots ovanstående är insektsobps av liten molekylstorlek med enkel produktion av rekombinanta proteiner, vilket gör dem favoritmål för strukturstudier och snabbbindande screening., Till exempel kan ligandscreening med OBPs möjliggöra identifiering av kemiska egenskaper för bättre bindning, såsom kedjelängd, molekylvolym, funktionella grupper och bond-ombildning. Dessa, i kombination med nya proteinstruktur förutsägelse metoder som används i utformningen av medicinska läkemedel och antikroppar, såsom homologi modellering, dynamik simuleringar och molekylär dockning, kan placera insekts OBPs i en favorit position över ORs som mål för utvecklingen av kontrollmedel i skadedjursbekämpning.
insekts Ors verkar mer specifikt inställd på luktämnen än OBPs., Den högre specificiteten som visas av ORs och risken för aktivering/hämning av specifika receptorer för ett visst beteende gör dessa proteiner som attraktiva mål för att manipulera skadedjursbeteenden. Genomförbarheten av hämning av antingen ORx / ORco-komplexet eller ORco av antagonister innefattar en lovande strategi för att störa insektspecifikt beteende, såsom parning via sexferomonreceptorer. Bristen på strukturell information är dock flaskhalsen när det gäller att använda insekts-eller insektsmedel som mål för semiokemiska aktivitetsprognoser., Tabell 1, 2 sammanfattar antalet OBPs och ORs som har identifierats hos insektsarter genom transkriptom (dvs RNA-seq) och genomsekvensering. De flesta insekter som studerats hittills har minst två gånger ORs än OBPs enligt genomstudier. Dessutom finns det en omfattande expansion av ORs i sociala insekter från Hymenoptera ordningen som honungsbiet A. mellifera med 170 ORs (Weinstock et al., 2006), och myrorna Solenopsis invicta och Cerapachys biroi med 400 och 506 ORs, respektive (Wurm et al., 2011; Oxley et al., 2014)., På samma sätt är eller expansionen också uppenbar i vissa jordbruksskadegörare, såsom den röda mjölbaggen T. castaneum med 265 ORs jämfört med 47 OBPs (Richards et al., 2008). Detta gör en krävande uppgift för målet eller valet tillsammans med svårigheten för det funktionella uttrycket av transmembranproteiner som ORs för att skärm ett stort antal ligander. En approximation av viktiga egenskaper i både OBP och ORs sammanfattas i tabell 3.
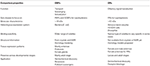
tabell 3., Ungefärlig jämförelse av insekts ORs och OBPs enligt egenskaper.
ytterligare perspektiv
den funktionella karakteriseringen av insekts-eller insektsområden samt deras bevisade roller i insekts olfaction har belyst känsligheten och specificiteten hos dessa insektspecifika proteiner. Dessa framsteg kommer att ytterligare förbättra deras genomförbarhet som skadedjursbekämpningsmål genom att förstå molekylär igenkänningsmekanismer och kombinatoriska interaktioner med OBP., Å andra sidan kommer den nuvarande massiva ansträngningen att identifiera och binda karakterisering av OBPs i flera viktiga insektsarter inom jordbruket att fortsätta och ge mer information om deras funktioner i insektsfysiologi. Således föreslår denna översyn som största fördel för OBPs över ORs, tillgängligheten av 3D-kristall-och NMR-strukturer, som med nedströmsmetoder, såsom homologimodellering (vid behov), molekylär dockning och molekyldynamik, skulle förfina sökningen av bioaktiva kemikalier., Detta sista som komplement till ligand affinitetsmätning kommer att påskynda studien av insektsobps som ska omprövas som målen för semiokemisk upptäckt och verktygen för att utforma superligander i skadedjursbekämpning.
utseendet och utvecklingen av insektsresistens hos skadeinsekter har lett till intensiv forskning om insekts olfaction och de mekanismer som är involverade för neural bearbetning. Det är väl etablerat att ett antal receptorer och enzymer i insekts CNS är målen för insektsresistensutveckling (Figur 1)., Det har visats att acetylkolinesteras (AChE) i löslig form ger resistens mot organiska fosfor och karbamatinsekticider, som fungerar som bioscavengers (Lee et al., 2015). På liknande sätt har flera insektsresistensmekanismer visats i bladlusen M. persicae, som involverar karboxylesteraser, natriumkanaler, γ-aminosmörsyra (GABA) och nAChR (bas et al., 2014)., Som viktiga komponenter i insekts periferin nervsystem och viktiga aktörer i insektsbeteenden representerar både insektsobps och ORs alternativa mål för identifiering av föreningar med semiokemisk aktivitet (eller agonistisk effekt) och verktyg för att utforma starka antagonister för att förbättra önskade beteendemässiga svar av insektsskadegörare och minska användningen av insekticider och efterföljande motstånd.
Författarbidrag
HV skrev avsnitt om OBP: s struktur och ORs, utvecklade tabeller och figur., J-JZ tänkt idén till översiktsartikeln, skrev huvudavsnittet, såsom introduktion, OBPs funktion och struktur samt eller-OBP jämförelse.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelser
författarna vill tacka FONDECYT 3170433., J-JZ är tacksam för det ekonomiska stödet från Northeast Normal University och Jilin University, Kina för hans sabbatiska ledighet att studera i Kina. Rothamsted Research får ett bidrag med ytterligare finansiering från bioteknik och biovetenskap (BBSRC), Storbritannien.
Gonzalez, F., Witzgall, P., och Walker, W. B. (2017). Antenntranskriptomer av tre tortricida moths avslöjar förmodade konserverade kemosensoriska receptorer för sociala och habitat olfaktoriska signaler. Sci. Rep.7, 1-12. doi: 10.,1038/srep41829
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Kaissling, K. E. (2013). Kinetik av olfaktoriska svar kan till stor del bero på luktreceptorinteraktionen och den luktdeaktivering som postulerats för fluxdetektorer. J. Comp. Physiol. Neuroetol. Sensorisk Neural Behav. Physiol. 199, 879–196. doi: 10.1007/s00359-013-0812-z
PubMed Abstrakt | CrossRef Full Text | Google Scholar