Design och konstruktion av randomiserade bibliotek för att analysera Cas9 PAM-inställningar
PAM bibliotek som innehåller randomiserade DNA-sekvenser omedelbart nedströms en DNA-sekvens som ett komplement till den distansen på en guide RNA var som genereras och används för att empiriskt bestämma PAM erkännande av Typ II Cas9 endonucleases (Fig. 1)., Med styr-rna-spacer-målsekvensen som fixeras tjänar de randomiserade baserna som ett substrat för direkt avläsning av Cas9 – endonukleaspam-specificiteten. Randomiserade sekvenser infördes i en plasmid DNA-vektor i PAM regionen av en protospacer mål sekvens som visar perfekt homologi till guide RNA spacer T1 (CGCUAAAGAGGAAGAGGACA). Två bibliotek ökar i storlek och komplexitet från fem randomiserade baspar (1,024 potentiella PAM kombinationer) till sju randomiserade baspar (16,384 potentiella Pam kombinationer) genererades., Randomisering av 5 bp-biblioteket infördes genom syntesen av en enda oligonukleotid innehållande fem slumpmässiga rester. Den enkelsträngade oligonukleotiden omvandlades till en dubbelsträngad mall med PCR (ytterligare fil 1: figur S1A), klonad till plasmidvektorn (ytterligare fil 1: figur S1B) och omvandlades till E. coli enligt beskrivningen i avsnittet metoder., För att säkerställa optimal slumpmässighet i 7 bp PAM-biblioteket reducerades bibliotekets storlek och komplexitet genom att syntetisera fyra oligonukleotider som var och en innehåller sex slumpmässiga rester plus en sjunde fast Rest bestående av G, C, A respektive T. Var och en av de fyra OLIGONUKLEOTIDERNA omvandlades separat till dubbelsträngat DNA, klonat till vektor pTZ57R/T enligt beskrivningen i Metodavsnittet och omvandlades till E. coli enligt beskrivningen för 5 bp-biblioteket., Efter transformation återfanns plasmid DNA och kombinerades från var och en av de fyra 6 bp PAM-biblioteken för att generera ett randomiserat 7 bp PAM-bibliotek bestående av 16,384 möjliga Pam-kombinationer. För båda biblioteken validerades inkorporering av slumpmässighet genom djup sekvensering; undersökning av nukleotidkompositionen vid varje position i PAM-regionen med hjälp av en positionsfrekvensmatris (PFM) (Metodavsnitt och ) (ytterligare fil 1: figur S2A och B)., Fördelningen och frekvensen för varje PAM-sekvens i 5 bp och 7 bp randomiserade PAM-biblioteket visas i ytterligare fil 1: figurerna S3 respektive S4.
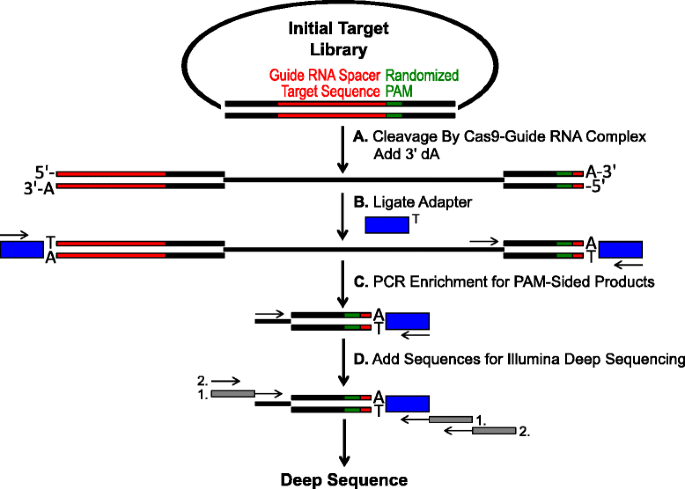
schematisk för identifiering av Pam-preferenser genom Cas9-klyvning in vitro. ett första plasmidbibliotek med randomiserad PAM (green box) klyvs med Cas9-komplex och 3′ da-överhäng läggs till. B adaptrar med 3′ dT överhäng (blue box) ligeras till båda ändarna av klyvprodukten., C Primers används för att berika för PAM-sidiga klyvda produkter av PCR., d Efter PCR-berikning, DNA-fragment, renas och Illumina kompatibel ankare och streckkoder är ”tailed-on” genom två omgångar av PCR (grå rutor) och Illumina djup analyserats
Analyseras Cas9 PAM-inställningar
Den randomiserade PAM bibliotek som beskrivs i föregående avsnitt utsattes för in vitro-rötning med olika koncentrationer av rekombinant Cas9 protein förladdad med guide RNA i syfte att analysen Cas9 endonuclease PAM önskemål på ett dosberoende sätt., Efter digestion med Cas9-guide rna ribonukleoprotein (RNP) komplex, Pam sekvenskombinationer från det randomiserade PAM-biblioteket som stödde klyvning fångades genom ligering adaptrar till de fria ändarna av de plasmida DNA-molekyler som klyvs av Cas9-guide RNA-komplexet (Fig. 1a och B). För att främja effektiv ligering och infångning av de klyvda ändarna modifierades den trubbiga dubbelsträngade DNA-skärningen som genererades av Cas9-endonukleaser för att innehålla en 3′ da-överhäng och Adaptrar ändrades för att innehålla en kompletterande 3′ dT-överhäng., För att generera tillräckliga mängder DNA för sekvensering förstärktes DNA-fragment som hyser PAM-sekvensen som stöder klyvning med hjälp av en primer i adaptern och en annan direkt intill PAM-regionen (Fig. 1c). De resulterande PCR förstärkta Cas9 Pam-biblioteken omvandlades till ampli-seq-mallar (Fig. 1D) och enkelläst djup sekvenseras från adaptern-sidan av amplicon., För att säkerställa tillräcklig täckning sekvenserades Cas9 PAM-biblioteken till ett djup minst fem gånger större än mångfalden i det ursprungliga randomiserade PAM-biblioteket (5,120 och 81,920 läser för 5 respektive 7 bp PAM randomiserade bibliotek). PAM-sekvenser identifierades från de resulterande sekvensdata genom att endast välja de läsningar som innehåller en perfekt 12 nt-sekvens match flankerar vardera sidan av 5 eller 7 nt PAM-sekvensen (beroende på det randomiserade PAM-biblioteket som används); fånga endast de PAM-sekvenser som härrör från perfekt Cas9-guide RNA-målplatsigenkänning och klyvning., För att kompensera för inneboende bias i de initiala randomiserade PAM-biblioteken normaliserades frekvensen för varje PAM-sekvens till dess frekvens i startbiblioteket. Eftersom analysen som beskrivs här direkt fångar Cas9 klyvbara PAM-sekvenser, användes probabilistisk modellering för att beräkna Pam-konsensusen för varje Cas9-protein. Detta åstadkoms genom att utvärdera sannolikheten för att hitta varje nukleotid (G, C, A eller T) vid varje position av PAM-sekvensen oberoende med hjälp av en positionsfrekvensmatris (PFM) (Metodavsnitt och )., De resulterande sannolikheterna visualiserades sedan som en webblogo .
för att undersöka benägenheten för falska positiva i analysen utelämnades tillsatsen av Cas9 RNP-komplex i digestionssteget (Fig. 1a) och analysen utfördes genom PCR-anrikningssteget (Fig. 1c). Som visas i ytterligare fil 1: figur S5A detekterades inga förstärkningsprodukter i frånvaro av Cas9-guide RNA-komplex. Således indikerar att förekomsten av falska positiva är låg och inte signifikant bidrar till resultaten av analysen.,
PAM preferenser av Streptococcus pyogenes och Streptococcus thermophilus (CRISPR3 och CRISPR1 system) Cas9 proteiner
för att bekräfta analysen, PAM preferenser av Streptococcus pyogenes (Spy) och Streptococcus thermophilus CRISPR3 (Sth3) Cas9 proteiner, vars PAM sekvens krav har tidigare rapporterats , undersöktes. In vitro-nyheter genomfördes med 1 µg (5.6 nM) av den 5 bp randomiserade PAM-biblioteket på två koncentrationer, 0,5 och 50 nM, förmonterade Spy eller Sth3 Cas9 protein, crRNA, och tracrRNA RNP komplex för 1 h på en 100 µL reaktion volym., Baserat på deras frekvens i 5 bp randomiserade PAM-biblioteket, Spy och Sth3 Cas9 PAM-sekvenser (NGG och NGGNG, respektive) var vid slutliga koncentrationer på 0.40 nM och 0.11 nM i matsmältningen, respektive. Medlemmar i det randomiserade PAM-biblioteket som innehöll PAM-sekvenser som stödde klyvning fångades och identifierades som beskrivs i föregående avsnitt. Som en negativ kontroll var det startande orenade randomiserade PAM-biblioteket föremål för sekvensering och PFM-analys tillsammans med de bibliotek som exponerades för Cas9 RNP-komplex., Som visas i Ytterligare fil 1: Figur S5B och C, ingen ordning inställningar finns i avsaknad av Cas9 RNP komplexa matsmältningen som framgår av en nära perfekt distribution av varje nukleotid på varje position på PAM i PFM bord och avsaknaden av informativa innehållet i WebLogo för kontroll. Detta är i stark constrast med Fig. 2a och b som visar sammansättningen av de sekvenser som härrör från bibliotek smälts med Spy och Sth3 Cas9 RNP komplex. Undersökning av PFM-härledda webbloggar (Fig., 2a och b) avslöjar också närvaron av de kanoniska PAM-preferenserna för Spionen och Sth3 Cas9-proteinerna, NGG respektive NGGNG. Även om Pam-preferenserna rapporterade för Spy och Sth3 Cas9-proteiner observeras i både 0.5 nM och 50 nM-digesterna, finns det en allmän breddning i specificitet under 50 nM-smältförhållandena. Detta är mest uppenbart vid position 2 för Spy Cas9-proteinet där frekvensen av en icke-kanonisk Rest ökar dramatiskt (Fig. 2a)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
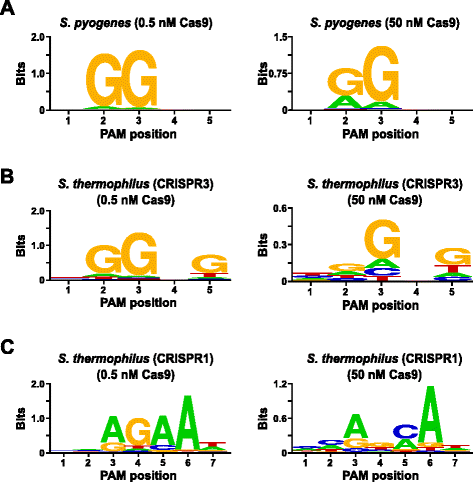
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., Frekvensen av nukleotider vid varje PAM position var självständigt beräknas med hjälp av en position frekvens matris (PFM) och plottas som en WebLogo
Ytterligare validering av analysen genomfördes genom att undersöka PAM preferenser för Streptococcus thermophilus CRISPR1 (Sth1) Cas9 protein vars PAM specificitet har rapporterats att utvidga ut till 7 bp . Med 1 µg (5.6 nM) 7 bp randomiserade PAM-biblioteket som mall, Sth1 Cas9-guide RNA digestions genomfördes i två koncentrationer, 0.5 nM 50 nM, RNP komplex som beskrivs ovan., Som kontroller användes spion-och Sth3 Cas9 RNP-komplex också för att smälta 7 bp randomiserade PAM-biblioteket men endast vid singel, 0.5 nM, RNP-komplex koncentration. Baserat på frekvensen i 7 bp randomiserade PAM-biblioteket var PAM-sekvenserna tidigare rapporterade för Sth1 (NNAGAAW), Spy (NGG) och Sth3 (NGGNG) vid slutliga koncentrationer av 0.01 nM, 0.22 nM respektive 0.05 nM., Som visas i Ytterligare fil 1: Figur S6A och B, PAM-inställningar för att Spy och Sth3 Cas9 proteiner som genereras med hjälp av den 7 bp bibliotek var nästan identiska med dem som produceras med 5 bp-biblioteket tillhandahåller ett starkt bevis för reproducerbarheten av analys. PAM preferenser för Sth1 Cas9 protein också matchade som tidigare rapporterats, NNAGAAW , på 0,5 nM Cas9-guide RNA komplexa koncentration (Fig. 2c)., Liknande Spy och Sth3 Cas9 proteiner, Sth1 Cas9 var kan klyva en mer varierad uppsättning av PAM sekvenser i de reaktioner som innehåller en högre koncentration av Cas9-guide RNA-komplex (50 nM), det mest slående var den markanta förlust av G rester krav på position 4 och nära lika preferens för en C-och En bp på position 5 (Fig. 2c). Detta resulterade i en annan PAM-konsensus än den som uppnåddes vid lägre koncentrationer.,
för Att pröva om PAM specificitet är oberoende av den typ av guide RNA, dubbelsidiga crRNA:tracrRNA eller sgRNA , Spy, Sth3, och Sth1 Cas9 PAM preferenser undersöktes också med hjälp av en binär Cas9 och sgRNA RNP komplex. Digestion utfördes vid en enda RNP-komplex koncentration av 0,5 nM och Pam preferensanalys utfördes som beskrivits ovan. Som visas i Ytterligare fil 1: Figur S7A, B, och C, PAM preferenser var nästan identiska oavsett vilken typ av guide RNA användas; antingen ett crRNA:tracrRNA duplex eller sgRNA., För att bekräfta att Pam-specificiteten inte påverkas starkt av sammansättningen av mål-DNA-eller spacersekvensen, var sekvensen på motsatt sida av 5 eller 7 bp randomiserade biblioteket riktad för klyvning med en annan spacer; T2-5 (UCUAGAUAGAUUACGAAUUUC) för 5 bp-biblioteket eller T2-7 (CCGGCGACGUUGGGUCAACU) för 7 bp-biblioteket. Spy och Sth3 Cas9 proteiner förladdad med sgRNAs inriktning T2-sekvens används för att förhöra 5 bp randomiserade PAM-biblioteket medan Sth1 Cas9-T2 sgRNA komplex användes för att smälta 7 bp randomiserade PAM-biblioteket., Pam preferenser analyserades som beskrivits ovan. PAM-inställningar för alla 3 Cas9 proteiner var nästan identisk oberoende av distansen och mål-DNA sekvens (fil 1: Figur S8A, B och C).
Identifiering av sgRNA och PAM preferenser för Brevibacillus laterosporus Cas9 protein
för Att empiriskt undersöka PAM preferenser för en Cas9 protein vars PAM var odefinierat, en uncharacterized Typ II-C CRISPR-Cas-lokus från Brevibacillus laterosporus stam SSP360D4 (Blat) identifierades genom att söka inre DuPont Pioneer databaser för Cas9 orthologues., Locus (cirka 4,5 kb) innehöll en cas9 gen som kan koda till 1 092 polypeptid, en CRISPR array bestående av sju upprepa-spacer enheter strax nedströms den cas9 gen och en tracrRNA kodning region som ligger uppströms cas9 gen med viss homologi till CRISPR mängd upprepningar (Fig. 3a). Repeat och distanslängd (36 och 30 bp, följaktligen) liknar andra typ II CRISPR-Cas-system med fem av de åtta upprepningarna innehållande 1 eller 2 bp-mutationer (Fig. 3b och Ytterligare fil 1: Figur S9)., Andra gener som normalt finns i en Typ II CRISPR-Cas-lokus var antingen trunkerad (cas1) eller saknas (Fig. 3a).
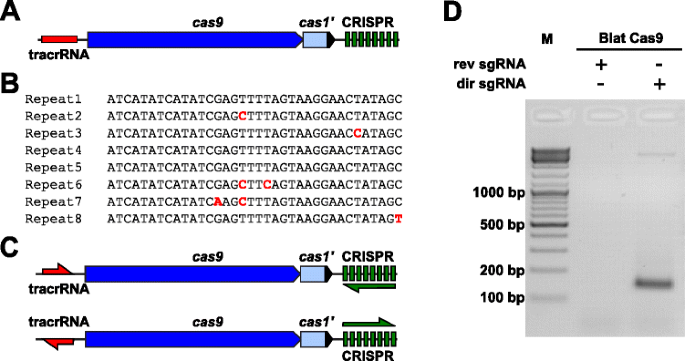
Identifiering av Typ II CRISPR-Cas-element i Brevibacillus laterosporus SSP360D4 CRISPR-Cas-systemet. en illustration av genomiskt DNA-region från Typ II CRISPR-Cas-systemet från Brevibacillus laterosporus SSP360D4. b-Jämförelse av Typ II CRISPR array upprepa sekvenser som identifierats i Brevibacillus laterosporus SSP360D4., c Den ”direkta” och ”omvänd” tracrRNA och CRISPR array transkriptionell scenarier för Typ II CRISPR-Cas-systemet från Brevibacillus laterosporus SSP360D4. d En agarosgel med reaktionsprodukter, som anger att endast den ”direkta’ sgRNA (dir sgRNA), men inte ”omvänd” sgRNA (rev sgRNA) stöd plasmid bibliotek klyvning i kombination med Cas9 endonuclease med ursprung från Brevibacillus laterosporus SSP360D4
guide RNA krav för Blat Cas9 protein bestämdes genom att generera två sgRNA varianter., Dessa varianter genererades att beakta både möjliga mening eller anti-sense uttryck scenarier av tracrRNA och CRISPR array (Fig. 3C) och används för att sondera vilket uttrycksscenario som stödde klyvningsaktiviteten hos Blat Cas9 i det randomiserade PAM-biblioteket. Single guide rna utformades genom att först identifiera gränserna för de förmodade tracrRNA-molekylerna genom att analysera regioner som delvis kompletterar 22 nt 5-terminalen av upprepningen (Anti-repeat)., Därefter, för att bestämma 3-änden av tracrRNA, användes möjliga sekundära strukturer och terminatorer för att förutsäga termineringsområdet i nedströmsfragmentet. Detta åstadkoms genom screening för närvaron av Rho-oberoende termineringssekvenser i DNA som omger antirepetitionen liknande den som beskrivs i Karvelis et al. , omvandla det omgivande DNA till RNA-sekvens och undersöka de resulterande strukturerna med hjälp av oförfalskad., Den resulterande sgRNAs utformades för att innehålla en T7 polymeras transkription initiering igenkänningssignal vid 5 ’slutet följt av en 20 nt mål igenkänningssekvens, 16 nt av crRNA upprepa, 4 nt själv fällbara hårnål slinga, och anti-upprepa sekvens som kompletterar upprepa regionen av crRNA följt av den återstående 3′ del av putative tracrna. SgRNA-varianten som innehåller en putativ tracrna transkriberas i samma riktning som cas9-genen (Fig. 3c) som kallas ”direkt” sgRNA, medan sgRNA innehåller tracrRNA transkriberas i motsatt riktning en ”omvänd” sgRNA., Femtio nM i Blat Cas9 sgRNA RNP komplexa, förinstallerade med antingen ”direkt” eller ”omvänd” sgRNAs respektive inkuberades med 1 µg (5.6 nM) 7 bp randomiserade PAM-biblioteket. Efter bibliotekets matsmältning och tillsats av 3’ dA överhäng ligerades adaptrar och klyvningsprodukter förstärktes PCR (Fig. 1). Analys av reaktionsprodukter genom agarosegelelektrofores visade att den ”direkta” sgRNA, men inte den ”omvända” sgRNA stödde plasmidbibliotekets klyvning (Fig. 3d). Sekvensen och den förväntade sekundära strukturen för ”direct” sgRNA visas i ytterligare fil 1: figur S10.,
Efter att bestämma en lämplig guide RNA för Blat Cas9, PAM identifiering utfördes på samma sätt som beskrivs ovan för att Spy, Sth3, och Sth1 Cas9 proteiner mot 7 bp randomiserade PAM-biblioteket med två koncentrationer, 0,5 och 50 nM, förmonterade Blat Cas9 ”direkt” sgRNA RNP komplex. Som visas i Fig. 4a, den PFM WebLogo PAM konsensus för Blat Cas9 protein under 0,5 nM smälta villkoren var NNNNCND (N = G, C, A, eller T; D = A, G, T) med en stark preferens för ett C i position 5 av PAM-sekvens., En måttlig preferens för en A observerades vid position 7 och små preferenser för en C eller T vid position 4 och G, C eller A över T vid position 6 noterades också vid noggrann granskning av PFM-tabellen (ytterligare fil 1: figur S11). Likaså att Spy, Sth3, och Sth1 Cas9 proteiner, PAM specificitet breddar som Cas9-sgRNA komplexa ökar koncentrationen. Detta är mest uppenbart vid position 5 där en större andel PAM-sekvenser innehållande en reststödsklyvning vid 50 nM jämfört med 0,5 nM-smältförhållandena.
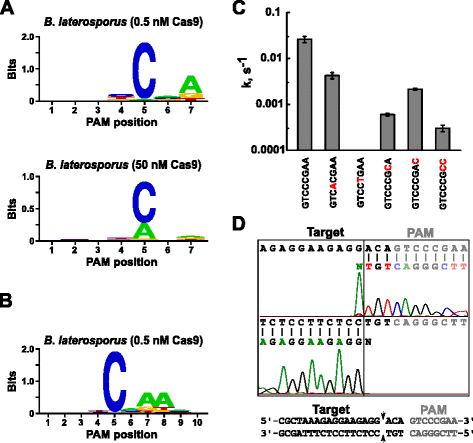
Pam-preferenser och klyvningspositioner för Brevibacillus laterosporus ssp360d4 (Blat) Cas9-enzym. Blat Cas9 PAM preferenser när 1 µg bibliotek DNA klyvs med 0,5 nM 50 nM Cas9-sgRNA komplex (a), ut på plats 10 av att flytta protospacer målet med 3 bp (b). Frekvensen av nukleotider vid varje PAM-position beräknades oberoende med hjälp av en positionsfrekvensmatris (PFM) och ritades som en webblogo ., c Klyvning priser av supercoiled plasmid DNA substrat som innehåller mutationer (visas i rött) i GTCCCGAA PAM-sekvens. Alla datapunkter är medelvärden från ≥3 oberoende experiment. Felstavar ges som S. D. D Run-off sekvensering från både sense och anti-sense riktningar plasmid DNA klyvs med Blat Cas9
eftersom Blat Cas9 kan acceptera någon bas i de tre första positionerna i sin PAM sekvens (Fig. 4A), distansen T1 skiftades med tre nukleotider i 5′ riktning för att tillåta Pam identifiering förlängas från 7 till 10 bp., De skiftade T1 spacer, T1-3 (AAACGCUAAAGAGGAAGAGG), införlivades i Blat ”direkt” sgRNA och PAM identifiering utförs som tidigare beskrivits för att Spy, Sth3, Sth1, och Blat Cas9 proteiner. Pam preferensanalys avslöjade Pam-specificiteten för Blat Cas9 kan utökas till position 8 där det finns en måttlig preferens för ytterligare A (Fig. 4b).
Pam specificitet för Blat Cas9 bekräftades genom att generera plasmider att innehålla mutationer i de mest konserverade rester av PAM (Fig. 4c)., Byte av C nukleotid på position 5 avskaffas plasmid DNA klyvning som bekräftar dess viktiga roll i Blat Cas9 PAM erkännande. Substitution av en nukleotider vid positionerna 7 och 8 signifikant reducerad (43× och 12×, respektive) klyvningshastigheten för supercoiled plasmid indikerar också vikten av dessa nukleotider i Blat Cas9 PAM erkännande.,
för Att identifiera DNA-mål klyvning positioner för Blat Cas9 protein, en plasmid som innehåller ett 20 bp regionen matchande distansen T1 följt av en PAM-sekvens, GTCCCGAA, som faller inom PAM konsensus för Blat Cas9, NNNNCNDD, togs fram och rötas med Blat Cas9-guide RNA ribonucleoprotein komplex. Direkt DNA-sekvensering användes för att bestämma ändarna av den linjära DNA-molekylen som genereras av Blat Cas9 RNP-komplexet. Sekvensen resultaten bekräftade att plasmid DNA klyvning uppstod i protospacer 3 nt 5 ” av PAM-sekvens (Fig., 4d) liknande den som observerades för att Spy, Sth3, och Sth1 Cas9 proteiner .
planta I genomet redigering använda Blat Cas9 och sgRNA
Följande belysning av sgRNA och PAM preferenser för Blat Cas9, majs optimerad Cas9 och sgRNA uttryck kassetter genererades i planta tester som tidigare beskrivits för S. pyogenes cas9 gen och sgRNA . Kort sagt var Blat cas9-genen majskodon optimerad och intron 2 av potatisst – LSI-genen infördes för att störa uttryck i E. coli och underlätta optimal Splitsning i planta (ytterligare fil 1: figur S12)., Kärnkraft lokalisering av Blat Cas9 protein i majs-celler underlättas genom tillsats av både aminosyror och carboxyl-terminal kärnkraft platser signaler, SV40 (MAPKKKRKV) och Agrobacterium tumefaciens VirD2 (KRPRDRHDGELGGRKRAR), respektive (fil 1: Figur S12). Den Blat cas9 gen var konstitutivt uttryckta i växternas celler genom att länka den optimerade cas9 till en majs Ubiquitin projektansvarig och pinII terminator i en plasmid DNA-vektor., För att ge ett effektivt sgRNA-uttryck i majsceller isolerades en promotor och terminator för majs U6 polymeras III (TTTTTTTTTTT) och smältes till 5-och 3-ändarna av en modifierad Blat sgRNA-kodning av DNA-sekvens (ytterligare fil 1: figur S13). Den modifierade Blat sgRNA innehöll två modifieringar från den som användes i in vitro-studierna; en t-g-förändring vid position 99 och en T-C-ändring vid position 157 i sgRNA (ytterligare fil 1: figur S13). Förändringarna infördes för att avlägsna potentiella för tidiga U6-polymeras III-termineringssignaler i Blat sgRNA., Förändringar där de infördes för att ha minimal inverkan på sekundärstrukturen hos sgRNA jämfört med den version som används i in vitro-studierna (data visas inte).
för Att exakt jämföra mutational effektivitet härrör från ofullständig icke-homolog end-anslutning (NHEJ) reparation av den dubbla DNA-strängbrott (DSBs) som härrör från Spy och Blat Cas9 klyvning, protospacer identiska dna-målet platser valdes ut genom att identifiera mål med Spy och Blat Cas9 kompatibel PAMs, NGGNCNDD., Identiska spacersekvenser valdes för Blat och Spy Cas9 genom att fånga 18 till 21 nt-sekvensen omedelbart uppströms PAM. För att säkerställa optimal U6 polymeras III uttryck och inte införa en obalans inom sgRNA spacer, alla målsekvenser valdes för att naturligt avsluta i en G vid deras 5′ slut. Mål identifierades och valdes ut i exon 1 och 4 av majsens fertilitetsgen Ms45 och i en region uppströms majsliguleless-1-genen.,
mutational aktivitet i Blat Cas9 i majs har granskats av biolistically omvandla 10 dagar gamla omogna majs embryon (IMEs) med DNA-vektorer som innehåller cas9 och sgRNA gener. Blat och motsvarande Spion Cas9 och sgRNA expressionsvektorer introducerades självständigt i majs Hi-typ II Imes genom partikelpistoltransformation som liknar den som beskrivs i . Eftersom partikelpistoltransformation kan vara mycket variabel, var en visuell markör DNA expression kassett, Ds-Röd, också co-levereras med Cas9 och sgRNA expression vektorer för att underlätta valet av jämnt transformerade Imes., Totalt utfördes tre transformationsreplikater på 60-90 Imes och 20-30 av de mest jämnt transformerade Imes från varje replikat skördades 3 dagar efter omvandling. Totalt genomiskt DNA extraherades och regionen kring målplatsen förstärktes av PCR och amplicons sekvenserade till ett läsdjup över 300,000. De resulterande läsningar undersöktes för närvaron av mutationer på den förväntade platsen för klyvning i jämförelse med kontroll experiment där sgRNA DNA uttryck kassett utelämnades från omvandlingen. Som visas i Fig., 5A, mutationer observerades vid den förväntade platsen för klyvning för Blat Cas9 med de vanligaste typerna av mutationer som enda baspar Infogningar eller deletioner. Liknande reparationsmönster observerades också för Spy Cas9-proteinet (ytterligare fil 1: figur S14 och ). Den mutational aktivitet för Blat Cas9 var stadig på två av de tre platser testade och överträffade att Spy Cas9 på Ms45 exon 4 mål webbplats med cirka 30 % (Fig. 5b).
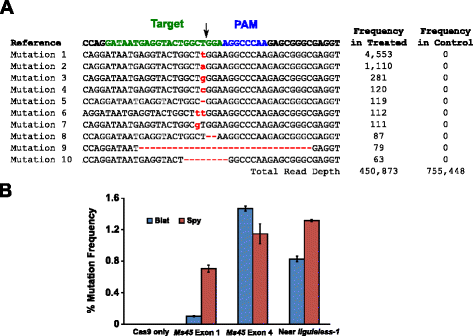
Brevibacillus laterosporus Cas9 främjar nhej-mutationer i majs. en Topp 10 mest vanliga typer av NHEJ mutationer som detekteras med Blat Cas9 i exon 4 av Ms45 gen. En svart pil indikerar den förväntade platsen för klyvning; mutationer markeras i rött; gemener teckensnitt indikerar en infogning; ’ – ’ indikerar en radering. b jämförelse av Spy och Blat Cas9 nhej mutation frekvenser vid tre protospacer identiska målområden i majs. Nhej-mutationer detekterades genom djup sekvensering 3 dagar efter omvandling., Fel barer representerar SEM, n = 3 partikel Pistol transformationer. Cas9 är endast den negativa kontrollen och representerar den genomsnittliga (över alla tre målplatser) bakgrundsfrekvensen för mutationer som härrör från PCR-förstärkning och sekvensering