Design og konstruktion af randomiserede biblioteker for at analysere Cas9 PAM preferences
PAM biblioteker, som indeholder randomiseret DNA-sekvenser umiddelbart efter af en DNA-sekvens, der er komplementær til den spacer af en guide RNA blev genereret, og som anvendes til empirisk at bestemme, PAM anerkendelse af Type II Cas9 endonucleases (Fig. 1)., Når guide RNA spacer-målsekvensen er fastgjort, tjener de randomiserede baser som et substrat til den direkte udlæsning af Cas9 endonuclease PAM-specificitet. Randomiserede sekvenser blev introduceret i en plasmid-DNA-vektor i PAM-regionen i en protospacer-målsekvens, der demonstrerede perfekt homologi til guide RNA-spacer T1 (cgcuaaagaggaagaggaca). To biblioteker vokser i størrelse og kompleksitet fra fem randomiserede basepar (1,024 potentielle PAM-kombinationer) til syv randomiserede basepar (16,384 potentielle PAM-kombinationer) blev genereret., Randomisering af 5 BP-biblioteket blev introduceret ved syntese af et enkelt oligonukleotid indeholdende fem tilfældige rester. Det enkeltstrengede oligonukleotid blev omdannet til en dobbeltstrenget skabelon ved PCR (yderligere fil 1: Figur S1A), klonet til plasmidvektoren (yderligere fil 1: Figur S1B) og omdannet til E. coli som beskrevet i afsnittet metoder., For at sikre optimal tilfældighed i 7 BP PAM-biblioteket blev bibliotekets størrelse og kompleksitet reduceret ved at syntetisere fire oligonukleotider, der hver indeholdt seks tilfældige rester plus en syvende fast rest omfattende henholdsvis G, C, A eller T. Hver af de fire oligonukleotider blev separat omdannet til dobbeltstrenget DNA, klonet til vektor pt .57r/t som beskrevet i Metodeafsnittet og omdannet til E. coli som beskrevet for 5 bp-biblioteket., Efter transformation blev plasmid-DNA genvundet og kombineret fra hvert af de fire 6 bp PAM-biblioteker for at generere et randomiseret 7 BP PAM-Bibliotek Omfattende 16.384 mulige PAM-kombinationer. For begge biblioteker blev inkorporering af tilfældighed valideret ved dyb sekventering; undersøgelse af nukleotidsammensætningen ved hver position i PAM-regionen ved hjælp af en positionsfrekvensmatri. (PFM) (Metodesektion og ) (yderligere fil 1: Figur S2A og B)., Fordelingen og hyppigheden af hver PAM sekvens i 5 bp og 7 bp randomiseret PAM bibliotek er vist i yderligere fil 1: tallene S3 og S4, henholdsvis.
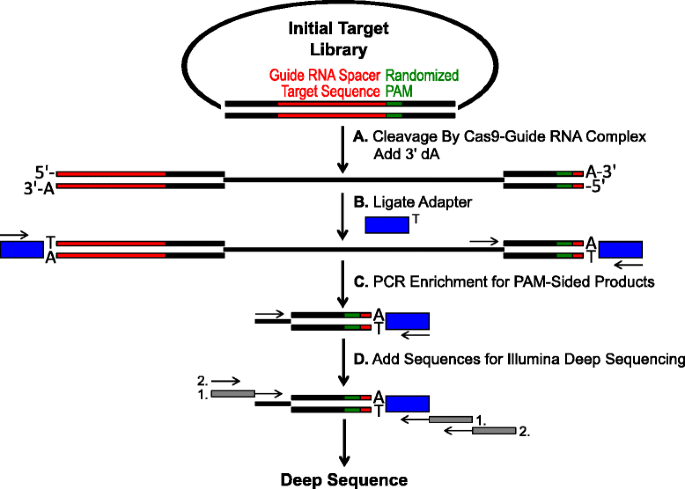
Skematisk til identifikation af PAM præferencer ved Cas9 spaltning in vitro. et indledende plasmidbibliotek med randomiseret PAM (grøn boks) spaltes med Cas9-kompleks, og 3′ dA-overhæng tilføjes. B-adaptere med 3 ‘ dt-overhæng (blå boks) er ligeret til begge ender af spaltningsproduktet., c primere anvendes til at berige for PAM-sidet spaltede produkter ved PCR., d Efter PCR berigelse, DNA fragmenter, der er renset og Illumina kompatibel ankre og stregkoder er ‘hale-på’ gennem to runder af PCR (grå kasser) og Illumina dybt sekventeret
Analysering Cas9 PAM preferences
De randomiserede PAM biblioteker, der er beskrevet i det foregående afsnit blev udsat til in vitro-fordøjelse med forskellige koncentrationer af rekombinant Cas9 protein præinstalleret med guide RNA for at analysen Cas9 endonuclease PAM-indstillinger i en dosisafhængig måde., Efter fordøjelse med Cas9-guide RNA ribonucleoprotein (RNP) komplekser, PAM sekvens kombinationer fra de randomiserede PAM-biblioteket, der støttede spaltning blev fanget af ligating adaptere til de frie ender af plasmid-DNA-molekyler spaltes af Cas9-guide RNA-komplekset (Fig. 1a og b). For at fremme effektiv ligering og indfangning af de spaltede ender blev det stumpede dobbeltstrengede DNA-snit genereret af Cas9-endonukleaser modificeret til at indeholde et 3′ dA-overhæng, og adaptere blev modificeret til at indeholde et komplementært 3′ dt-overhæng., For at generere tilstrækkelige mængder DNA til sekventering blev DNA-fragmenter, der huser PAM-sekvensen, der understøtter spaltning, PCR forstærket under anvendelse af en primer i adapteren og en anden direkte ved siden af PAM-regionen (fig. 1c). De resulterende PCR-forstærkede Cas9 PAM-biblioteker blev konverteret til ampli-se. – skabeloner (fig. 1d) og enkeltlæs dybt sekventeret fra adaptersiden af ampliconen., For at sikre tilstrækkelig dækning blev Cas9 PAM-bibliotekerne sekventeret til en dybde, der var mindst fem gange større end mangfoldigheden i det oprindelige randomiserede PAM-bibliotek (5,120 og 81,920 læser for henholdsvis 5 og 7 BP PAM randomiserede biblioteker). PAM-sekvenser blev identificeret fra den viste sekvens data ved kun at vælge dem, der læser, der indeholder en perfekt 12 nt sekvens matcher flankerende hver side af 5 eller 7 nt PAM sekvens (afhængigt af de randomiserede PAM-biblioteket bruges); at fange kun de PAM-sekvenser som følge af perfekt Cas9-guide RNA-mål site anerkendelse og spaltning., For at kompensere for iboende bias i de indledende randomiserede PAM-biblioteker blev frekvensen af hver PAM-sekvens normaliseret til dens frekvens i startbiblioteket. Da analysen beskrevet her direkte fanger Cas9-spaltelige PAM-sekvenser, blev probabilistisk modellering brugt til at beregne PAM-konsensus for hvert Cas9-protein. Dette blev opnået ved at evaluere sandsynligheden for at finde hvert nukleotid (G, C, A eller T) ved hver position af PAM-sekvensen uafhængigt ved hjælp af en positionsfrekvensmatri. (PFM) (Metodesektion og)., De resulterende sandsynligheder blev derefter visualiseret som Enbleblogo .
for at undersøge tilbøjeligheden til falske positiver i assayet blev tilsætningen af Cas9 RNP-komplekser i fordøjelsestrinnet udeladt (fig. 1a), og analysen blev udført gennem PCR-berigelsestrinnet (fig. 1c). Som vist i yderligere fil 1: Figur S5A blev der ikke påvist forstærkningsprodukter i fraværet af Cas9-guide RNA-komplekser. Således indikerer, at forekomsten af falske positiver er lav og ikke bidrager væsentligt til resultaterne af analysen.,
PAM præferencer af Streptococcus pyogenes og Streptococcus thermophilus (CRISPR3 og CRISPR1 systemer) Cas9 proteiner
for at validere analysen, PAM præferencer af Streptococcus pyogenes (Spion) og Streptococcus thermophilus CRISPR3 (Sth3) Cas9 proteiner, hvis PAM sekvens krav har tidligere været rapporteret , blev undersøgt. In vitro fordøjer blev gennemført med 1 µg (5.6 nM) af de 5 bp randomiseret PAM-biblioteket i to koncentrationer, 0,5 og 50 nM, af pre-samlet Spion eller Sth3 Cas9 protein, crRNA, og tracrRNA RNP komplekser i 1 time i en 100 µL volumen reaktion., Baseret på deres frekvens i 5 bp randomiseret PAM-biblioteket, Spion og Sth3 Cas9 PAM-sekvenser (NGG og NGGNG, henholdsvis) var i endelige koncentrationer på 0,40 nM og 0,11 nM i fordøjelsen, hhv. Medlemmer af det randomiserede PAM-bibliotek, der indeholdt PAM-sekvenser, der understøttede spaltning, blev fanget og identificeret som beskrevet i det foregående afsnit. Som en negativ kontrol, start ukløvet randomiseret PAM-biblioteket blev genstand for sekventering og PFM analyse sammen med de biblioteker, der udsættes for Cas9 RNP-komplekser., Som vist i Ekstra fil 1: Figur S5B og C, ingen rækkefølge præferencer, der eksisterer i mangel af Cas9 RNP komplekse fordøjelsen så tydeligt, som en nær perfekt fordeling af nucleotiderne på hver position af PAM i PFM bordet og manglende informative indhold i WebLogo for kontrol. Dette er i skarp kontrast med Fig. 2a og b, der illustrerer sammensætningen af sekvenser afledt af biblioteker fordøjet med Spy og Sth3 Cas9 RNP komplekser. Undersøgelse af PFM-afledte Weeblogos (Fig., 2a og b) viser også tilstedeværelsen af den kanoniske PAM præferencer for Spy og Sth3 Cas9 proteiner, NGG og NGGNG , hhv. Selv om PAM præferencer, der indberettes for Spy og Sth3 Cas9 proteiner er observeret i både 0,5 nM 50 nM fordøjer, der er en generel udvidelse i specificitet under 50 nM fordøje betingelser. Dette er mest tydeligt i position 2 For Spy Cas9-proteinet, hvor hyppigheden af en ikke-kanonisk a-rest øges dramatisk (fig. 2a)., For Sth3, all PAM positions exhibit a marked decrease in specificity as a result of increasing the RNP complex concentration (Fig. 2b).
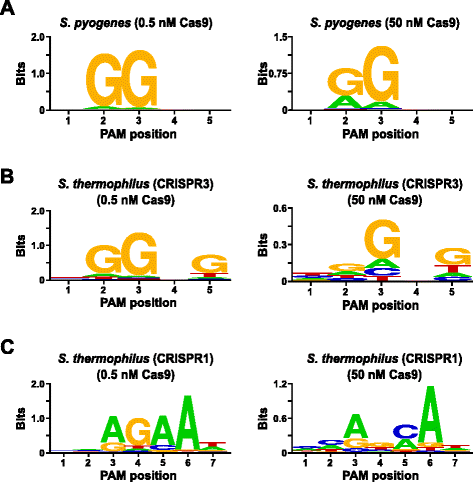
PAM preferences for S. pyogenes (a), S. thermophilus CRISPR3 (b), and S. thermophilus CRISPR1 (c) Cas9 proteins., Hyppigheden af nukleotider på hver PAM position uafhængigt beregnet ved hjælp af en position frekvens matrix (PFM) og afbildet som en WebLogo
en Yderligere validering af analysen blev gennemført ved at undersøge PAM præferencer for Streptococcus thermophilus CRISPR1 (Sth1) Cas9 protein, hvis PAM specificitet er blevet rapporteret til at strække sig til 7 bp . Ved anvendelse af 1 µg (5.6 nM) af det 7 bp randomiserede PAM-bibliotek som skabelon blev Sth1 Cas9-guide RNA-fordøjelser udført ved to koncentrationer, 0.5 nM og 50 nM, af RNP-kompleks som beskrevet ovenfor., Som kontroller blev Spy-og Sth3 Cas9 RNP-komplekser også brugt til at fordøje det 7 bp randomiserede PAM-bibliotek, men kun ved enkelt, 0.5 nM, RNP-kompleks koncentration. Baseret på den frekvens i 7 bp randomiseret PAM-biblioteket, PAM-sekvenser, der tidligere er rapporteret for Sth1 (NNAGAAW), Spy (NGG), og Sth3 (NGGNG) var i endelige koncentrationer på 0,01 nM, 0.22 nM, og 0,05 nM hhv., Som vist i yderligere fil 1: Figur S6A og B, PAM-præferencerne for Spy-og Sth3 Cas9-proteiner genereret ved hjælp af 7 BP-biblioteket var næsten identiske med dem, der blev produceret med 5 bp-biblioteket, der leverede stærkt bevis for reproducerbarheden af assayet. PAM-præferencerne for Sth1 Cas9-proteinet matchede også tæt den tidligere rapporterede, NNAGAA., ved 0, 5 nM Cas9-guide RNA-kompleks koncentration (fig. 2c)., Lignende for at Udspionere og Sth3 Cas9 proteiner, Sth1 Cas9 var i stand til at spalte et mere mangfoldigt sæt af PAM-sekvenser i de reaktioner, der indeholder en højere koncentration af Cas9-guide RNA-kompleks (50 nM), er det mest slående var den markante tab af G rest krav på position 4 og nær samme præference for et C og Et kogepunkt på position 5 (Fig. 2c). Dette resulterede i en anden PAM-konsensus end den, der blev opnået ved lavere koncentrationer.,
for At undersøge, om PAM specificitet er uafhængig af den type guide, RNA, dupleksudskrevne crRNA:tracrRNA eller sgRNA , Spion, Sth3, og Sth1 Cas9 PAM præferencer blev også undersøgt ved hjælp af en binær Cas9 og sgRNA RNP komplekse. Fordøjelsen blev udført ved en enkelt RNP kompleks koncentration på 0,5 nM og PAM præferenceanalyse blev udført som beskrevet ovenfor. Som vist i Ekstra fil 1: Figur S7A, B, og C, PAM præferencer var næsten identiske, uanset type guide, RNA, der anvendes; enten en crRNA:tracrRNA duplex eller sgRNA., Hertil kommer, at bekræfte, at PAM specificitet er ikke i høj grad påvirket af sammensætningen af mål-DNA eller spacer sekvens, den sekvens, på den modsatte side af 5 eller 7 bp randomiseret bibliotek var målrettet for spaltning med en anden spacer; T2-5 (UCUAGAUAGAUUACGAAUUC) til 5 bp bibliotek eller T2-7 (CCGGCGACGUUGGGUCAACU) for de 7 bp bibliotek. Spion og Sth3 Cas9 proteiner præinstalleret med sgRNAs rettet mod T2 sekvens blev brugt til at afhøre 5 bp randomiseret PAM-biblioteket, mens Sth1 Cas9-T2 sgRNA komplekser blev brugt til at fordøje den 7 bp randomiseret PAM-biblioteket., PAM præferencer blev analyseret som beskrevet ovenfor. PAM-indstillinger for alle 3 Cas9 proteiner var næsten identiske, uanset spacer og mål-DNA sekvens (Ekstra fil 1: Figur S8A, B og C).
Identifikation af sgRNA og PAM præferencer for Brevibacillus laterosporus Cas9 protein
empirisk At undersøge PAM præferencer for en Cas9 protein, hvis PAM var udefineret, en uncharacterized Type II-C CRISPR-Cas-locus fra Brevibacillus laterosporus stamme SSP360D4 (Blat) blev identificeret ved at søge indre DuPont Pioneer databaser for Cas9 orthologues., Locus (ca 4,5 kb) indeholdt en cas9 gen stand til indkodning af et 1,092 polypeptid, en CRISPR array bestående af syv repeat-spacer enheder lige neden for cas9 gen og en tracrRNA kodning region, som ligger oven for cas9 gen med delvis homologi til den CRISPR array gentagelser (Fig. 3a). Gentag og spacer længde (36 og 30 bp, i overensstemmelse hermed) svarer til anden Type II CRISPR-Cas-systemer med fem af de otte gentagelser, der indeholder 1 eller 2 bp mutationer (Fig. 3b og yderligere fil 1: Figur S9)., Andre gener typisk findes i en type II CRISPR-Cas locus blev enten afkortet (cas1) eller mangler (fig. 3a).
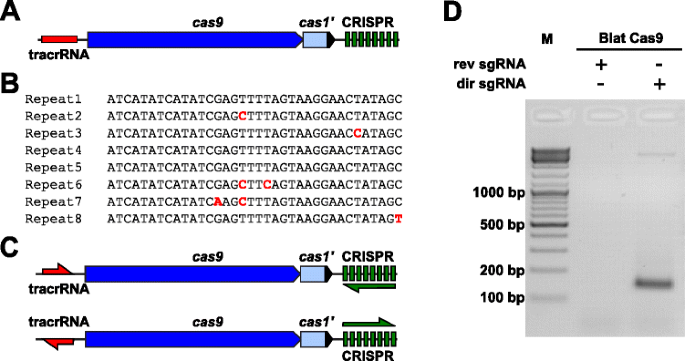
Identifikation af Type II CRISPR-Cas elementer i Brevibacillus laterosporus SSP360D4 CRISPR-Cas-system. en illustration af den genomiske DNA-region fra type II CRISPR-Cas-systemet fra Brevibacillus laterosporus ssp360d4. B sammenligning af type II CRISPR array repeat sekvenser identificeret i Brevibacillus laterosporus ssp360d4., c ‘direkte’ og ‘omvendt’ tracrRNA og CRISPR array transkriptionel scenarier for Type II CRISPR-Cas-system fra Brevibacillus laterosporus SSP360D4. d En agarose-gel med reaktionsprodukter, der angiver, at kun den ‘direkte’ sgRNA (dir sgRNA), men ikke ‘omvendt’ sgRNA (rev sgRNA) støtte plasmid bibliotek spaltning i kombination med Cas9 endonuklease, der stammer fra Brevibacillus laterosporus SSP360D4
The guide RNA krav til Blat Cas9 protein blev bestemt ved at generere to sgRNA varianter., Disse varianter blev genereret for at tage højde for både mulige sanse-eller anti-sense-ekspressionsscenarier af tracrRNA – og CRISPR-arrayet (fig . 3c) og bruges til at undersøge hvilket ekspressionsscenarie understøttet spaltningsaktivitet af Blat Cas9 i det randomiserede PAM-bibliotek. Enkelt guide-RNA’ er blev designet ved først at identificere grænserne for de formodede tracrRNA-molekyler ved at analysere regioner, der delvis var komplementære til 22 NT 5 ‘ – terminalen for gentagelsen (anti-gentagelse)., For at bestemme 3 ‘ – enden af tracrRNA blev mulige sekundære strukturer og terminatorer brugt til at forudsige termineringsområdet i nedstrøms fragmentet. Dette blev opnået ved screening for tilstedeværelsen af Rho-uafhængige-lignende termineringssekvenser i DNA ‘ et omkring anti-repeat svarende til det, der er beskrevet i Karvelis et al. , omdanne det omgivende DNA til RNA-sekvens og undersøge de resulterende strukturer ved hjælp af UNAfold ., Den resulterende sgRNAs var designet til at indeholde en T7-polymerase transkription indledningen anerkendelse signal på 5? ende efterfulgt af en 20 nt mål anerkendelse sekvens, 16 nt af crRNA gentage, 4 nt self-folde-hairpin loop, og anti-gentag sekvensen er komplementær til at gentage region crRNA efterfulgt af de resterende 3’ del af den formodede tracrRNA. SgRNA-varianten, der indeholder et formodet tracrRNA transkriberet i samme retning som cas9-genet (fig . 3c) betegnes ‘direkte’ sgRNA, mens sgRNA indeholdende tracrRNA transskriberet i modsat retning en ‘omvendt’ sgRNA., Halvtreds nM Blat Cas9 sgRNA RNP-kompleks, forudindlæst med henholdsvis ‘direkte’ eller ‘omvendt’ sgrna ‘ er, blev inkuberet med 1 µg (5.6 nM) af det 7 bp randomiserede PAM-bibliotek. Efter biblioteksfordøjelse og tilsætning af 3 ‘ dA-overhæng blev adaptere ligeret, og spaltningsprodukter blev PCR-forstærket (fig. 1). Analyse af reaktionsprodukter ved agarosegelelektroforese afslørede, at den’ direkte ‘sgRNA, men ikke den’ omvendte ‘ sgRNA understøttede plasmidbibliotekspaltning (fig. 3d). Sekvensen og den forudsagte sekundære struktur af ‘direkte’ sgRNA er vist i yderligere fil 1: Figur S10.,
Efter fastsættelsen af en passende guide RNA til Blat Cas9, PAM identifikation blev udført på samme måde som beskrevet ovenfor for Spion, Sth3, og Sth1 Cas9 proteiner mod 7 bp randomiseret PAM-biblioteket med to koncentrationer, 0,5 og 50 nM, af pre-samlet Blat Cas9 ‘direkte’ sgRNA RNP komplekse. Som vist i Fig. 4a, den PFM WebLogo PAM konsensus for Blat Cas9 protein under 0,5 nM fordøje betingelser var NNNNCND (N = G, C, A, T, D = A, G eller T), med en stærk præference for en C på position 5 af PAM-sekvens., En moderat præference for En blev observeret ved position 7 og lille præferencer for en C eller T på position 4 og G, C eller over En T på position 6 blev også bemærket, når du nøje undersøger, PFM tabel (flere filer 1: Figur S11). På samme måde som Spy -, Sth3-og Sth1 Cas9-proteiner udvides PAM-specificiteten, når Cas9-sgRNA-komplekskoncentrationen stiger. Dette er mest tydeligt i position 5, hvor en større andel af PAM-sekvenser indeholdende en a-reststøttespaltning ved 50 nM sammenlignet med 0,5 nM-fordøjelsesbetingelserne.
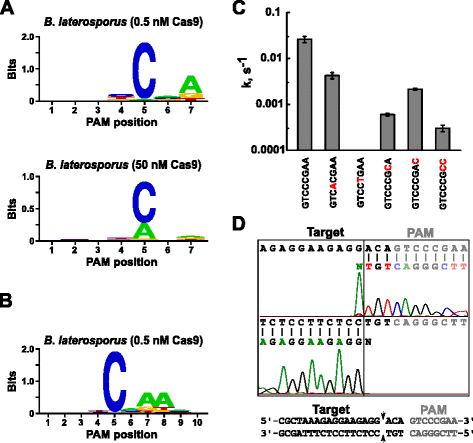
PAM præferencer og spaltning holdninger Brevibacillus laterosporus SSP360D4 (Blat) Cas9 enzym. Blat Cas9 PAM præferencer, når 1 µg bibliotek DNA er kløvet med 0,5 nM eller 50 nM Cas9-sgRNA kompleks (en), forlænget ud til position 10 ved at flytte protospacer mål på 3 bp (b). Frekvensen af nukleotider ved hver PAM-position blev uafhængigt beregnet ved hjælp af en positionsfrekvensmatri. (PFM) og afbildet som Enbleblogo., c Spaltningshastigheder af supercoiled plasmid DNA substrater indeholdende mutationer (vist i rødt) i GTCCCGAA PAM sekvens. Alle datapunkter er middelværdier fra 3 3 uafhængige eksperimenter. Fejl barer er givet, da S. D. Run-off sekventering fra både fornuft og anti-fornuft retninger af plasmid DNA spaltes med Blat Cas9
Da Blat Cas9 kan acceptere nogen base i de første tre positioner af sine PAM rækkefølge (Fig. 4a) blev afstandsstykket T1 forskudt med tre nukleotider i 5′ – retningen for at tillade PAM-identifikation at blive udvidet fra 7 til 10 bp., De forskudte T1 spacer, T1-3 (AAACGCUAAAGAGGAAGAGG), blev indarbejdet i Blat ‘direkte’ sgRNA og PAM identifikation blev udført, som beskrevet tidligere Spion, Sth3, Sth1, og Blat Cas9 proteiner. PAM-præferenceanalyse afslørede, at PAM-specificiteten for Blat Cas9 kan udvides til position 8, hvor der er en moderat præference for et yderligere A (fig. 4b).
PAM specificitet for Blat Cas9 blev bekræftet ved at generere plasmider til at indeholde mutationer i de mest velbevarede rester af PAM (Fig. 4c)., Udskiftning af C-nukleotid ved position 5 afskaffet plasmid-DNA-spaltning, der bekræfter dens nøglerolle i Blat Cas9 PAM-anerkendelse. Udskiftning af nukleotider i positionerne 7 og 8 reducerede signifikant (henholdsvis 43 and og 12 respectively) spaltningshastigheden af supercoiled plasmid, hvilket også indikerer betydningen af disse nukleotider i Blat Cas9 PAM-genkendelse.,
for At identificere DNA-mål spaltning holdninger til Blat Cas9 protein, et plasmid, der indeholder en 20 bp regionen matchende spacer T1 efterfulgt af et PAM-sekvens, GTCCCGAA, der falder inden for PAM konsensus for Blat Cas9, NNNNCNDD, der blev genereret og fordøjet med Blat Cas9-guide RNA ribonucleoprotein komplekse. Direkte DNA-sekventering blev brugt til at bestemme enderne af det lineære DNA-molekyle genereret af Blat Cas9 RNP-komplekset. Sekvensresultaterne bekræftede, at plasmid-DNA-spaltning forekom i protospaceren 3 NT 5′ i PAM-sekvensen (fig., 4d) svarende til den, der observeres for Spy -, Sth3-og Sth1 Cas9-proteiner .
I planta genom-redigering med Blat Cas9 og sgRNA
Følgende udredning af sgRNA og PAM præferencer for Blat Cas9, majs optimeret Cas9 og sgRNA udtryk kassetter blev genereret i planta test, som tidligere beskrevet for S. pyogenes cas9 gen og sgRNA . Kort fortalt blev Blat cas9-genet optimeret med majskodon, og intron 2 af kartoffel ST-LSI-genet blev indsat for at forstyrre ekspression i E. coli og lette optimal splejsning i planta (yderligere fil 1: Figur S12)., Nuklear lokalisering af Blat Cas9 proteinet i majs celler blev lettet ved tilsætning af både aminosyrer og carboxylgrupper-terminal nukleare steder signaler, SV40 (MAPKKKRKV) og Agrobacterium tumefaciens VirD2 (KRPRDRHDGELGGRKRAR), henholdsvis (Ekstra fil 1: Figur S12). Blat cas9-genet blev konstitutivt udtrykt i planteceller ved at forbinde den optimerede cas9 med en majs UBI .uitin-promotor og pinII-terminator i en plasmid-DNA-vektor., For at give effektiv sgRNA udtryk i majs-celler, en majs U6-polymerase III arrangøren og terminator (TTTTTTTT) blev isoleret og smeltet til 5′ – og 3′ – ender i en modificeret Blat sgRNA kodning DNA-sekvens, henholdsvis (Ekstra fil 1: Figur S13). Den modificerede Blat sgRNA indeholdt to modifikationer fra den, der blev anvendt i in vitro-undersøgelserne; en T til g-ændring ved position 99 og en T til C-ændring ved position 157 i sgRNA (yderligere fil 1: Figur S13). Ændringerne blev introduceret for at fjerne potentielle for tidlige U6-polymerase III-termineringssignaler i Blat sgRNA., Ændringer, der er indført for at have minimal indvirkning på den sekundære struktur af sgRNA sammenlignet med den version, der blev anvendt i in vitro-undersøgelserne (data ikke vist).
Til præcist at sammenligne mutational effektivitet som følge af den ufuldkomne ikke-homologe ende-at deltage (NHEJ) reparation af DNA-dobbelt-strand pauser (DSBs) som følge af Spion-og Blat Cas9 spaltning, protospacer identiske genomisk mål steder blev udvalgt ved at udpege mål med Spion og Blat Cas9 kompatibel PAMs, NGGNCNDD., Identiske afstandssekvenser blev valgt til Blat og Spy Cas9 ved at fange 18 til 21 NT-sekvensen umiddelbart opstrøms for PAM. For at sikre optimal U6-polymerase III-ekspression og ikke indføre en uoverensstemmelse inden for sgRNA-afstandsstykket blev alle målsekvenser valgt til naturligt at afslutte i en g ved deres 5′ ende. Mål blev identificeret og udvalgt i e .on 1 og 4 af majsfrugtbarhedsgenet Ms45 og i en region opstrøms for majs liguleless-1 genet.,
mutational aktivitet af Blat Cas9 i majs blev gennemgået af biolistically at omdanne 10 dage gamle umodne majs embryoner (ime ‘ er) med DNA-vektorer der indeholder cas9 og sgRNA gener. Blat og de tilsvarende Spion Cas9 og sgRNA ekspressionsvektorer blev uafhængigt introduceret i majs Hi-type II IME ‘ er ved partikelpistoltransformation svarende til den, der er beskrevet i . Da partikelpistoltransformation kan være meget variabel, blev en visuel markør DNA-ekspressionskassette, Ds-Red, også leveret sammen med Cas9-og sgRNA-ekspressionsvektorerne for at hjælpe med valget af jævnt transformerede IME ‘ er., I alt blev tre transformationsreplikater udført på 60-90 IME ‘er, og 20-30 af de mest jævnt transformerede IME’ er fra hver replikat blev høstet 3 dage efter transformation. Total genomisk DNA blev ekstraheret, og regionen omkring målstedet blev forstærket ved PCR og ampliconer sekventeret til en læsedybde på over 300.000. De resulterende læsninger blev undersøgt for tilstedeværelsen af mutationer på det forventede spaltningssted ved sammenligning med kontrolforsøg, hvor sgRNA DNA-ekspressionskassetten blev udeladt fra transformationen. Som vist i Fig., 5a, mutationer blev observeret på det forventede sted for spaltning for Blat Cas9, hvor de mest udbredte typer mutationer var Indsætninger eller deletioner af enkelt basepar. Lignende reparationsmønstre blev også observeret for Spy Cas9-proteinet (yderligere fil 1: Figur S14 og ). Den mutational aktivitet for Blat Cas9 var robust i to af de tre lokaliteter, testet og oversteg Spion Cas9 på Ms45 exon 4 mål site med omkring 30 % (Fig. 5b).
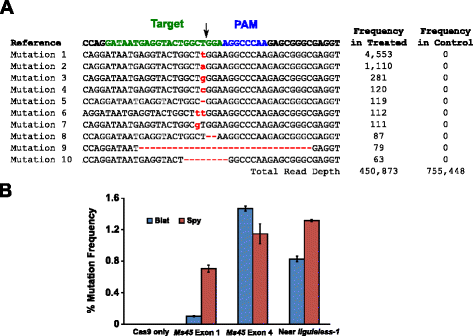
Brevibacillus laterosporus Cas9 fremmer NHEJ mutationer i majs. en top 10 mest udbredte typer af NHEJ mutationer påvist med Blat Cas9 i e .on 4 af ms45 genet. En sort pil angiver det forventede spaltningssted; mutationer fremhæves med rødt; små bogstaver angiver en indsættelse; ‘ – ‘ angiver en sletning. B sammenligning af spion-og Blat Cas9 NHEJ-mutationsfrekvenser ved tre protospacer-identiske målsteder i majs. NHEJ-mutationer blev påvist ved dyb sekventering 3 dage efter transformation., Fejl barer repræsenterer SEM, n = 3 partikel pistol transformationer. Cas9 kun er de negative kontrol-og repræsenterer gennemsnittet (på tværs af alle tre mål steder) baggrund hyppigheden af mutationer, som følge af PCR-amplifikation og sekventering