aunque los compuestos volátiles con actividad agonista se han examinado contra ORs, una sustancia química específica con fuerte efecto agonista sobre los mosquitos, 2-(4-etil-5-(piridin-3-Il)-4H-1,2,4-triazol-3-iltio)-N-(4-etilfenil)acetamida (VUAA1), ha abierto el campo de la investigación (Jones et al., 2011; Taylor et al., 2012). Estudios posteriores de Taylor et al., (2012) proporcionaron evidencia de productos químicos derivados de VUAA1, como VUAA4, capaces de aumentar su efecto agonista en 10 veces en ORco de A. gambiae, H. virescens y harpegnathos saltator. Curiosamente, los autores informaron que cualquier cambio en los sustituyentes de la amida causará una pérdida completa de la actividad agonista. Esto proporciona información útil sobre los requisitos estructurales de los agonistas y la relación estructura-actividad entre los análogos de VUAA y las RUP. Finalmente, a pesar de la actividad agonista mejorada de los productos químicos VUAA, su peso molecular relativamente alto (367.47 g mol−1 para VUAA1) vs., los agonistas volátiles, como bombykol (238.42 g mol-1), hacen que una entrega Volátil directa de VUAA no sea factible. Con esto en mente, la búsqueda de análogos estructurales más pequeños representa un foco interesante de investigación.
antagonismo en la subunidad ORco
junto con el estudio de análogos relacionados con VUAA que pueden actuar como agonistas fuertes, el bloqueo de ORco por antagonistas también ha surgido para guiar el diseño semioquímico y pesticida., Por lo tanto, un análogo estructural de VUAA1, VU0183254 (2-(4-etil-5-furan-2-Il-4h-triazol-3-ilsulfanil)-1-fenotiazina-10-Il-etanona), se informó que inhibe la respuesta ORco, actuando como modulador alostérico en A. gambiae e interrumpiendo el reconocimiento de agonistas como el eugenol por el complejo OR65/ORco (Jones et al., 2012). Otros análogos estructurales de VUAA también han sido reportados como antagonistas. Un ejemplo son los compuestos de triazolotioacetamida n -, 2 sustituidos OLC3 y OLC12 que interrumpen la respuesta ORco de manera similar en C. quinquefasciatus, A. gambiae, D. melanogaster y O., nubilalis, sugiriendo un sitio de unión conservado en ORco (Chen y Luetje, 2012). Considerando la inhibición de ORco como una estrategia prometedora para interrumpir el comportamiento de los insectos, parece que los esfuerzos posteriores deberían apuntar a los compuestos con menor peso molecular que los antagonistas derivados de VUAA. Por ejemplo, se informó que OX1a (232 g mol−1), triptamina (160,22 g mol−1) y cinamato de isopropilo (190,24 g mol−1) tenían efecto antagonista sobre ORco (Chen y Luetje, 2013, 2014; Tsitoura et al., 2015) con aproximadamente la mitad o menos de peso molecular que VUAA1., Sin embargo, el uso futuro de estos antagonistas debe ser estudiado cuidadosamente, ya que el bloqueo del ORco conservado puede afectar no solo a los insectos dañinos, sino también a los beneficiosos.
Además del efecto antagonista sondeado in vitro, la evidencia a nivel conductual apoya la idea de que los análogos estructurales de las feromonas pueden funcionar como antagonistas. Por ejemplo, Sellanes et al., (2010) reportaron la inhibición de la respuesta sexual en los Criptoblabos gnidiella de la polilla melosa cuando los análogos estructurales, formiato (Z)-9-tetradecenilo y formiato (Z)-11-hexadecenilo, se agregaron a la feromona sexual sintética, (Z)-11-hexadecenal y (Z)-13-octadecenal, en pruebas de túnel de viento. Este efecto antagonista de la feromona fue corroborado más tarde en ensayos de campo, donde el atrapamiento de los machos de C. gnidiella disminuyó en un patrón dosis-dependiente. El antagonismo de las feromonas también se ha reportado para B. mandarina, un ancestro de B. mori (Daimon et al., 2012)., Sus hallazgos corroboran bombykol como la feromona sexual, y bombykal y acetato de bombykyl como antagonistas, lo que inhibió fuertemente la atracción de los hombres en el campo a la feromona sexual bombykol. Más recientemente, la evidencia del antagonismo de la feromona fue divulgada para la polilla del hocico Herpetogramma submarginale. Cuando se añadió (Z)-13-hexadecenol a su feromona sexual, acetato de (Z)-13-hexadecenilo, disminuyó significativamente el número de machos capturados en el campo (Yan et al., 2015)., El antagonismo de las feromonas parece basarse en las diferencias en el grupo funcional químico, como alcoholes, aldehídos y ésteres, dependiendo de la especie de insecto. Sin embargo, el efecto antagonista de estos análogos estructurales puede no deberse a la inhibición de ORco, sino a la especificidad de ORx frente a antagonistas. Un estudio reciente sugiere que el OR16 de Helicoverpa armigera es capaz de reconocer específicamente el antagonista de la feromona, (Z)-11-hexadecenol (Chang H. et al., 2017). Los autores apoyaron el papel específico de OR16 considerando que H., las hembras de armigera emiten el compuesto antagonista junto con su feromona sexual ((Z)-11-hexadecenal Y (Z)-9-hexadecenal) como estrategia para evitar el apareamiento no óptimo con machos inmaduros. Sobresalientemente, cuando el OR16 fue derribado por la técnica de edición del genoma CRISPR / Cas9 y los machos de H. armigera fueron probados por Electrofisiología y ensayos conductuales, no se registró respuesta EAG y los machos intentaron aparearse con Hembras inmaduras.
Receptores Olfativos vs, Proteínas de unión: Pros y Contras para el manejo de plagas de insectos
para el caso de las OBPs, la especificidad del ligando y los mecanismos de las OBPs representan aspectos controvertidos, lo que parece fuertemente dependiente de los métodos utilizados para la medición de la afinidad del ligando. Por ejemplo, se ha informado de que los PBPs, como los de P. xylostella y eogystia hippophaecolus de las polillas, pueden unirse tanto a los componentes de feromonas sexuales como a los análogos (Sun et al., 2013a; Hu et al., 2018). Esto sugiere que los actores descendentes, como los ORs, podrían mejorar la especificidad y la sensibilidad de la recepción de odorantes., Evidencia reciente apoya que la co-expresión de PBPs y PRs puede aumentar la sensibilidad hacia feromonas. Por ejemplo, se utilizaron múltiples combinaciones de PR1-4 y PBP1-4 para probar su respuesta a los componentes de feromonas sexuales de la polilla Chilo supressalis (Chang et al., 2015). Los autores encontraron un aumento significativo en la sensibilidad de la respuesta hacia (Z)-11-hexadecenal cuando PR4 y PR6 se coexpresaron con PBP4., Aunque la interacción de estas proteínas podría surgir un nuevo nivel de investigación como objetivos de control de plagas, el diferente emparejamiento de PRs y PBPs arrojó luces sobre la complejidad del sistema olfativo en insectos, haciendo que el enfoque sea una tarea difícil para un gran conjunto de compuestos y proteínas para probar. A pesar de lo anterior, los OBP de insectos son de pequeño tamaño molecular con fácil producción de proteínas recombinantes, lo que los convierte en objetivos favoritos para estudios estructurales y detección de unión rápida., Por ejemplo, el cribado de ligandos con OBPs podría permitir la identificación de propiedades químicas para una mejor unión, como la longitud de la cadena, el volumen molecular, los grupos funcionales y la insaturación del enlace. Estos, combinados con los nuevos métodos de predicción de la estructura proteica utilizados en el diseño de medicamentos y anticuerpos, como el modelado de homología, las simulaciones dinámicas y el acoplamiento molecular, podrían colocar a los OBP de insectos en una posición preferida sobre los Or como objetivos para el desarrollo de agentes de control en el manejo de plagas.
Los or de insectos parecen estar más específicamente sintonizados con los odorantes que los OBP., La mayor especificidad mostrada por las ORs y la posibilidad de activación/inhibición de receptores específicos para un comportamiento dado hacen que estas proteínas sean objetivos atractivos para manipular comportamientos de plagas. La viabilidad de la inhibición del complejo ORx / ORco u ORco por antagonistas comprende una estrategia prometedora para interrumpir el comportamiento específico del insecto, como el apareamiento a través de receptores de feromonas sexuales. Sin embargo, la falta de información estructural es el cuello de botella en el uso de ORs de insectos como objetivos para las predicciones de actividad semioquímica., Las tablas 1, 2 Resumen El número de OBPs y ORs que se han identificado en especies de insectos por transcriptoma (es decir, ARN-seq) y secuenciación del genoma. La mayoría de los insectos estudiados hasta ahora tienen por lo menos dos veces ORs que OBPs según estudios del genoma. Además, hay una extensa expansión de ORs en insectos sociales del orden Himenóptero como la abeja melífera A. mellifera con 170 ORs (Weinstock et al., 2006), y las hormigas Solenopsis invicta y Cerapachys biroi con 400 y 506 ORs, respectivamente (Wurm et al., 2011; Oxley et al., 2014)., Del mismo modo, la expansión de la OR también es evidente en algunas plagas agrícolas, como el escarabajo de la harina roja T. castaneum con 265 Or en comparación con 47 OBPs (Richards et al., 2008). Esto hace una tarea exigente para el blanco o la selección junto con la dificultad para la expresión funcional de proteínas transmembrana como ORs con el fin de cribar un gran número de ligandos. En la Tabla 3 se resume una aproximación de las propiedades importantes tanto en los OBP como en los Or.
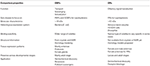
la Tabla 3., Comparación aproximada de ORs de insectos y OBP según propiedades.
más perspectivas
la caracterización funcional de las Or de insectos, así como sus funciones probadas en el olfato de insectos, han arrojado luz sobre la sensibilidad y especificidad de estas proteínas específicas de insectos. Estos avances mejorarán aún más su viabilidad como objetivos de control de plagas mediante la comprensión de los mecanismos de reconocimiento molecular y las interacciones combinatorias con los PAB., Por otro lado, el esfuerzo masivo Actual en la identificación y caracterización de unión de OBPs en varias especies de insectos agrícolas importantes continuará y proporcionará más información sobre sus funciones en la fisiología de los insectos. Por lo tanto, esta revisión propone como principal ventaja para los OBP sobre los ORs, la disponibilidad de estructuras de cristal 3D y RMN, que con enfoques descendentes, como el modelado de homología (cuando sea necesario), el acoplamiento molecular y la dinámica molecular, refinarían la búsqueda de productos químicos bioactivos., Este último en complemento con la medición de afinidad de ligandos acelerará el estudio de los OBP de insectos para ser reconsiderados como los objetivos para el descubrimiento semioquímico y las herramientas para diseñar superligandos en el manejo del control de plagas.
la aparición y el desarrollo de resistencia a insecticidas en plagas de insectos han llevado a la investigación intensiva sobre el olfato de insectos y los mecanismos que intervienen para el procesamiento neural. Está bien establecido que una serie de receptores y enzimas en los NAFTALENOS clorados de los insectos son los objetivos para el desarrollo de resistencia a los insecticidas (Figura 1)., Se ha demostrado que la acetilcolinesterasa (AChE) en forma soluble proporciona la resistencia a los insecticidas organofosforados y carbamatos, actuando como bioscavengers (Lee et al., 2015). De manera similar, se han demostrado múltiples mecanismos de resistencia a insecticidas en el áfido M. persicae, que involucran carboxilesterasas, canales de sodio, ácido γ-aminobutírico (GABA) y nAChR (Bass et al., 2014)., Como componentes importantes en el sistema nervioso periférico de los insectos y actores clave en el comportamiento de los insectos, tanto los OBP como los Or representan objetivos alternativos para la identificación de compuestos con actividad semioquímica (o efecto agonista) y herramientas para diseñar antagonistas fuertes para mejorar las respuestas de comportamiento deseadas de las plagas de insectos y reducir el uso de insecticidas y la resistencia posterior.
contribuciones de los autores
HV escribió secciones sobre la estructura de OBP y ORs, desarrolló tablas y figuras., J-JZ concibió la idea para el artículo de revisión, escribió la sección principal, como introducción, función y estructura OBPs, así como comparación OR-OBP.
Declaración de conflicto de intereses
los autores declaran que la investigación se realizó en ausencia de relaciones comerciales o financieras que pudieran ser interpretadas como un potencial conflicto de intereses.
Agradecimientos
Los autores agradecen a FONDECYT 3170433., J-JZ está agradecido por el apoyo financiero de la Northeast Normal University y la Universidad Jilin, China, para su licencia sabática para estudiar en China. Rothamsted Research recibe una subvención con financiación adicional de Biotechnology and Biological Sciences (BBSRC), Reino Unido.
Gonzalez, F., Witzgall, P., and Walker, W. B. (2017). Los transcriptomas antenales de tres polillas tortrídicas revelan supuestos receptores quimiosensoriales conservados para señales olfativas sociales y de hábitat. Sci. El representante. 7, 1-12. doi: 10.,1038/srep41829
PubMed Abstract | CrossRef Full Text/Google Scholar
Kaissling, K. E. (2013). La cinética de las respuestas olfativas podría depender en gran medida de la interacción odorante-receptor y la desactivación odorante postulada para los detectores de flujo. J. Comp. Physiol. Neuroetol. Comportamiento Neural Sensorial. Physiol. 199, 879–196. doi: 10.1007 / s00359-013-0812-z
resumen de PubMed | texto completo de CrossRef / Google Scholar