bien que des composés volatils ayant une activité agoniste aient été testés contre les SRO, un produit chimique spécifique ayant un fort effet agoniste sur les moustiques, le 2-(4-Ethyl-5-(pyridin-3-yl)-4h-1,2,4-triazol-3-ylthio)-n-(4-ethylphenyl)acetamide (VUAA1), a ouvert le champ de la recherche (Jones et al., 2011; Taylor et coll., 2012). Études ultérieures de Taylor et al., (2012) ont fourni des preuves de produits chimiques dérivés de VUAA1, tels que VUAA4, capables d’augmenter son effet agoniste de 10 fois sur L’ORco d’A. gambiae, H. virescens et harpegnathos saltator. Fait intéressant, les auteurs ont rapporté que tout changement sur les substituants amides entraînera une perte complète de l’activité agoniste. Cela donne des informations utiles sur les exigences structurelles des agonistes et la relation structure-activité entre les analogues de la VUAA et les SRO. Enfin, malgré l’activité agoniste accrue des produits chimiques VUAA, son poids moléculaire relativement élevé (367,47 g mol−1 pour VUAA1) vs., les agonistes volatils, tels que le bombykol (238,42 g mol−1), rendent impossible une administration volatile directe de VUAA. Dans cet esprit, la recherche d’analogues structurels plus petits représente un centre de recherche intéressant.
antagonisme sur la sous-unité ORco
parallèlement à l’étude des analogues de la VUAA qui peuvent agir comme agonistes puissants, le blocage de L’ORco par les antagonistes est également apparu pour guider la conception des produits semi-chimiques et des pesticides., Ainsi, un analogue structurel de VUAA1, VU0183254 (2 – (4-éthyl-5-furane-2-yl-4h-triazol-3-ylsulfanyl)-1-phénothiazine-10-YL-éthanone), a été rapporté pour inhiber la réponse ORco, agissant comme modulateur allostérique chez A. gambiae et perturbant la reconnaissance d’agonistes tels que l’eugénol par le complexe OR65/ORco (Jones et al., 2012). D’autres analogues structuraux de la VUAA ont également été rapportés comme antagonistes. Un exemple est le triazolothioacétamide N -, 2-substitué composés OLC3 et OLC12 qui perturbe la réponse ORco d’une manière similaire chez C. quinquefasciatus, A. gambiae, D. melanogaster, et O., nubilalis, suggérant un site de liaison conservé à ORco (Chen et Luetje, 2012). Considérant L’inhibition de L’ORco comme une stratégie prometteuse pour perturber les comportements des insectes, il semble que les efforts ultérieurs devraient viser les composés ayant un poids moléculaire inférieur à celui des antagonistes dérivés de la VUAA. Par exemple, L’OX1a (232 g mol−1), la tryptamine (160,22 g mol−1) et le cinnamate d’isopropyle (190,24 g mol−1) auraient un effet antagoniste sur L’ORco (Chen et Luetje, 2013, 2014; Tsitoura et al., 2015) avec environ la moitié ou moins de poids moléculaire que VUAA1., Néanmoins, l’utilisation future de ces antagonistes doit être étudiée attentivement, car le blocage de L’ORco conservé peut affecter non seulement les insectes nuisibles, mais aussi les insectes bénéfiques.
outre l’effet antagoniste étudié in vitro, les preuves au niveau comportemental soutiennent l’idée que les analogues structurels des phéromones peuvent fonctionner comme antagonistes. Par exemple, Sellanes et al., (2010) ont signalé l’inhibition de la réponse sexuelle chez la teigne du miellat Cryptoblabes gnidiella lorsque les analogues structuraux, Le formiate de (Z)-9-tétradécényle et le formiate de (z)-11-hexadécényle, ont été ajoutés à la phéromone sexuelle synthétique, le (Z)-11-hexadécénal et le (Z)-13-octadécénal, lors d’essais en soufflerie. Cet effet antagoniste des phéromones a ensuite été corroboré dans des essais sur le terrain, où le piégeage des mâles de C. gnidiella a diminué selon un schéma dose-dépendant. L’antagonisme des phéromones a également été rapporté pour B. mandarina, un ancêtre de B. mori (Daimon et al., 2012)., Leurs résultats corroborent le bombykol comme phéromone sexuelle, et le bombykal et l’acétate de bombykyle comme antagonistes, ce qui inhibe fortement l’attraction des mâles sur le terrain pour la phéromone sexuelle bombykol. Plus récemment, des signes d’antagonisme des phéromones ont été rapportés chez le papillon Herpetogramma submarginale. Lorsque le (Z)-13-hexadécénol a été ajouté à sa phéromone sexuelle, l’acétate de (z) – 13-hexadécényle a considérablement diminué le nombre de mâles capturés au champ (Yan et al., 2015)., L’antagonisme des phéromones semble basé sur les différences de groupe fonctionnel chimique tels que les alcools, les aldéhydes et les esters selon les espèces d’insectes. Néanmoins, l’effet antagoniste de ces analogues structuraux pourrait ne pas être dû à L’inhibition de L’ORco mais à la spécificité de L’ORx aux antagonistes. Une étude récente suggère que L’OR16 de Helicoverpa armigera est capable de reconnaître spécifiquement l’antagoniste des phéromones, le (Z)-11-hexadécénol (Chang H. et al., 2017). Les auteurs ont appuyé le rôle spécifique de L’OR16 considérant que H., les femelles d’armigera émettent le composé antagoniste avec sa phéromone sexuelle ((Z)-11-hexadécenal et (Z)-9-hexadécenal) comme stratégie pour éviter l’accouplement non optimal avec les mâles immatures. Fait remarquable, lorsque L’OR16 a été abattu par la technique D’édition du génome CRISPR / Cas9 et que les mâles de H. armigera ont été testés par électrophysiologie et tests comportementaux, aucune réponse EAG n’a été enregistrée et les mâles ont essayé de s’accoupler avec des femelles immatures.
récepteurs odorants vs., Protéines de liaison: avantages et inconvénients pour la lutte contre les insectes nuisibles
dans le cas des OBP, la spécificité et les mécanismes des OBP représentent des aspects controversés, qui semblent fortement dépendants des méthodes utilisées pour mesurer l’affinité des ligands. Par exemple, il a été rapporté que les PBP, tels que ceux de P. xylostella et Eogystia hippophaecolus, peuvent lier à la fois les composants et les analogues des phéromones sexuelles (Sun et al., 2013a; Hu et coll., 2018). Cela suggère que les acteurs en aval tels que les SRO pourraient améliorer la spécificité et la sensibilité de la réception des odeurs., Des preuves récentes soutiennent que la co-expression des PBP et des PRs peut augmenter la sensibilité aux phéromones. Par exemple, plusieurs combinaisons de PR1-4 et de PBP1-4 ont été utilisées pour tester leur réponse aux composants des phéromones sexuelles du papillon Chilo suppressalis (Chang et al., 2015). Les auteurs ont constaté une augmentation significative de la sensibilité de la réponse à (Z)-11-hexadécénal lorsque PR4 et PR6 étaient Co-exprimés avec PBP4., Bien que l’interaction de ces protéines puisse susciter un nouveau niveau de recherche en tant que cibles de lutte antiparasitaire, l’appariement différent des Pr et des PBP jette des lumières sur la complexité du système olfactif chez les insectes, ce qui rend l’approche difficile à tester pour un large ensemble de composés et de protéines. Malgré ce qui précède, les insectes OBP sont de petite taille moléculaire avec une production facile de protéines recombinantes, ce qui en fait des cibles préférées pour les études structurelles et le criblage de liaison rapide., Par exemple, le criblage de ligands avec OBPs pourrait permettre l’identification de propriétés chimiques pour une meilleure liaison, telles que la longueur de la chaîne, le volume moléculaire, les groupes fonctionnels et l’insaturation des liaisons. Celles-ci, combinées aux nouvelles méthodes de prédiction de la structure des protéines utilisées dans la conception de médicaments et d’anticorps, telles que la modélisation de l’homologie, les simulations dynamiques et l’amarrage moléculaire, pourraient placer les OBP des insectes dans une position préférée par rapport aux ORs en tant que cibles pour le développement d’agents de lutte antiparasitaire.
Les ORs des insectes semblent plus spécifiquement adaptés aux odeurs que les OBP., La spécificité plus élevée démontrée par les SRO et la possibilité d’activation/inhibition de récepteurs spécifiques pour un comportement donné font de ces protéines des cibles attrayantes pour manipuler les comportements des ravageurs. La faisabilité de l’inhibition du complexe ORx/ORco ou ORco par des antagonistes comprend une stratégie prometteuse pour perturber le comportement spécifique des insectes, tel que l’accouplement via les récepteurs des phéromones sexuelles. Cependant, le manque d’informations structurelles est le goulot d’étranglement dans l’utilisation des SRO d’insectes comme cibles pour les prévisions d’activité sémiochimique., Les tableaux 1 et 2 résument le nombre d’OBP et D’or qui ont été identifiés chez les espèces d’insectes par transcriptome (c.-À-D. ARN-seq) et séquençage du génome. La plupart des insectes étudiés jusqu’à présent ont au moins deux fois ORs que OBP selon les études du génome. De plus, il y a une expansion considérable des SRO chez les insectes sociaux de l’Ordre des hyménoptères tels que L’abeille mellifère A. mellifera avec 170 SRO (Weinstock et al., 2006), et les fourmis Solenopsis invicta et Cerapachys biroi avec 400 et 506 Rup, respectivement (Wurm et coll., 2011; Oxley et coll., 2014)., De même, l’expansion de la RO est également évidente chez certains ravageurs agricoles, tels que le coléoptère de la farine rouge t. castaneum avec 265 ro comparativement à 47 PBO (Richards et al., 2008). Cela rend une tâche exigeante pour la cible ou la sélection ainsi que la difficulté pour l’expression fonctionnelle des protéines transmembranaires telles que les SRO afin de filtrer un grand nombre de ligands. Une approximation des propriétés importantes des OBP et des ORs est résumée dans le tableau 3.
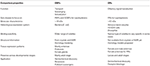
le Tableau 3., Comparaison approximative des insectes ORs et OBP selon les propriétés.
autres Perspectives
La caractérisation fonctionnelle des SRO des insectes ainsi que leur rôle avéré dans l’olfaction des insectes ont mis en lumière la sensibilité et la spécificité de ces protéines spécifiques aux insectes. Ces progrès amélioreront encore leur faisabilité en tant que cibles de lutte antiparasitaire grâce à la compréhension des mécanismes de reconnaissance moléculaire et des interactions combinatoires avec les OBP., D’autre part, l’effort massif actuel dans l’identification et la caractérisation de liaison des OBP chez plusieurs espèces d’insectes agricoles importantes se poursuivra et fournira plus d’informations sur leurs fonctions dans la physiologie des insectes. Ainsi, cette revue propose comme principal avantage pour les OBP par rapport aux ORs, la disponibilité de structures cristallines 3D et RMN, qui avec des approches en aval, telles que la modélisation d’homologie (si nécessaire), l’amarrage moléculaire et la dynamique moléculaire, affineraient la recherche de produits chimiques bioactifs., Ce dernier complément à la mesure de l’affinité des ligands accélérera l’étude des OBP des insectes à reconsidérer comme cibles de découverte sémiochimique et comme outils pour concevoir des super-ligands dans la lutte antiparasitaire.
L’apparition et le développement de la résistance aux insecticides chez les insectes nuisibles ont conduit à la recherche intensive sur l’olfaction des insectes et les mécanismes impliqués dans le traitement neuronal. Il est bien établi qu’un certain nombre de récepteurs et d’enzymes dans le SNC des insectes sont les cibles du développement de la résistance aux insecticides (Figure 1)., Il a été démontré que l’acétylcholinestérase (AChE) sous forme soluble fournit la résistance aux insecticides organo-phosphore et carbamate, agissant comme bioscavengers (Lee et al., 2015). De même, de multiples mécanismes de résistance aux insecticides ont été démontrés chez le puceron M. persicae, impliquant des carboxylestérases, des canaux sodiques, de l’acide γ-aminobutyrique (GABA) et du nAChR (Bass et al., 2014)., En tant que composants importants du système nerveux périphérique des insectes et acteurs clés dans les comportements des insectes, les OBP et les or représentent des cibles alternatives pour l’identification de composés ayant une activité semi-chimique (ou un effet agoniste) et des outils pour concevoir des antagonistes puissants pour améliorer les réponses comportementales souhaitées des insectes nuisibles et réduire l’utilisation
contributions des auteurs
HV a écrit des sections sur la structure de L’OBP et les ORs, développé des tableaux et des figures., J-JZ a conçu l’idée de l’article de revue, a écrit la section principale, telle que l’introduction, La fonction et la structure OBPs ainsi que la comparaison OR-OBP.
déclaration de conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de toute relation commerciale ou financière pouvant être interprétée comme un conflit d’intérêts potentiel.
Remerciements
Les auteurs tiennent à remercier FONDECYT 3170433., J-JZ est reconnaissant pour le soutien financier de L’Université normale du Nord-Est et de L’Université de Jilin, en Chine, pour son congé sabbatique pour étudier en Chine. Rothamsted Research reçoit une subvention avec un financement supplémentaire de Biotechnology and Biological Sciences (BBSRC), Royaume-Uni.
Gonzalez, F., Witzgall, P., et Walker, W. B. (2017). Les transcriptomes antennaires de trois papillons tortricides révèlent des récepteurs chimiosensoriels conservés pour les signaux olfactifs sociaux et d’habitat. Sci. Rép. 7, 1-12. doi: 10.,1038/srep41829
PubMed Abstract | CrossRef Texte Intégral | Google Scholar
Kaissling, K. E. (2013). La cinétique des réponses olfactives pourrait largement dépendre de l’interaction odorant-récepteur et de la désactivation odorante postulée pour les détecteurs de flux. J. Comp. Physiol. Neuroéthol. Comportement Neuronal Sensoriel. Physiol. 199, 879–196. doi: 10.1007/s00359-013-0812-z
PubMed Abstract | CrossRef Texte Intégral | Google Scholar