Obwohl flüchtige Verbindungen mit Agonistenaktivität gegen ORs, eine spezifische Chemikalie mit starker Agonistenwirkung auf Mücken, 2-(4-Ethyl-5-(Pyridin-3-yl)-4H-1,2,4-Triazol-3-ylthio)-N-(4-ethylphenyl)Acetamid (VUAA1), untersucht wurden, wurde das Gebiet der forschung (Jones et al., 2011; Taylor et al., 2012). Spätere Studien von Taylor et al., (2012) lieferte Beweise für von VUAA1 abgeleitete Chemikalien wie VUAA4, die in der Lage waren, ihre agonistische Wirkung auf ORco von A. gambiae, H. virescens und Harpegnathos saltator um das Zehnfache zu erhöhen. Interessanterweise berichteten die Autoren, dass jede Änderung der Amidsubstituenten zu einem vollständigen Verlust der Agonistenaktivität führt. Dies liefert hilfreiche Einblicke in die strukturellen Anforderungen von Agonisten und die Struktur-Aktivitäts-Beziehung zwischen VUAA-Analoga und ORs. Schließlich, trotz der erhöhten Agonistenaktivität von VUAA−Chemikalien, sein relativ hohes Molekulargewicht (367.47 g mol-1 für VUAA1) vs., flüchtige Agonisten, wie Bombykol (238,42 g mol−1), macht eine direkte flüchtige Abgabe von VUAA etwas nicht machbar. Vor diesem Hintergrund stellt die Suche nach kleineren Strukturanaloga einen interessanten Forschungsschwerpunkt dar.
Antagonismus der ORco-Untereinheit
Zusammen mit der Untersuchung von VUAA-bezogenen Analoga, die als starke Agonisten wirken können, hat sich auch die Blockade von ORco durch Antagonisten herausgestellt, um Semiochemikalien und Pestiziddesign zu leiten., So wurde berichtet, dass ein strukturelles Analogon von VUAA1, VU0183254 (2-(4-Ethyl-5-furan-2-yl-4H-triazol-3-ylsulfanyl)-1-phenothiazin-10-yl-ethanon), die ORco-Reaktion hemmt, als allosterischer Modulator in A. gambiae wirkt und die Erkennung von Agonisten wie Eugenol durch den Komplex OR65/ORco stört (Jones et al., 2012). Andere VUAA-strukturelle Analoga wurden ebenfalls als Antagonisten berichtet. Ein Beispiel sind die N -, 2-substituierten Triazolothioacetamidverbindungen OLC3 und OLC12, die die ORco-Reaktion in ähnlicher Weise in C. quinquefasciatus, A. gambiae, D. melanogaster und O stören., nubilalis, was auf eine konservierte Bindungsstelle in ORco hindeutet (Chen und Luetje, 2012). Betrachtet man die Hemmung von ORco als eine vielversprechende Strategie, um das Verhalten von Insekten zu stören, scheint es, dass nachfolgende Bemühungen auf die Verbindungen mit einem niedrigeren Molekulargewicht als VUAA-abgeleitete Antagonisten abzielen sollten. Es wurde beispielsweise berichtet, dass OX1a (232 g mol−1), Tryptamin (160,22 g mol−1) und Isopropylcinnamat (190,24 g mol−1) eine antagonistische Wirkung auf ORco haben (Chen und Luetje, 2013, 2014; Tsitoura et al., 2015) mit etwa der Hälfte oder weniger Molekulargewicht als VUAA1., Dennoch sollte die zukünftige Verwendung dieser Antagonisten sorgfältig untersucht werden, da die Blockade des konservierten ORco nicht nur schädliche, sondern auch nützliche Insekten betreffen kann.
Neben dem in vitro untersuchten Antagonisteneffekt unterstützen die Beweise auf Verhaltensebene die Idee, dass strukturelle Analoga von Pheromonen als Antagonisten fungieren können. Für Beispiel, Sellanes et al., (2010) berichtete über die Hemmung der sexuellen Reaktion in der Honigtaumotte Cryptoblabes gnidiella, wenn die strukturellen Analoga, (Z)-9-Tetradecenylformiat und (Z)-11-Hexadezienylformiat, zu synthetischem Sexualpheromon hinzugefügt wurden, (Z)-11-Hexadezial und (Z)-13-Octadecenal, in Windkanaltests. Dieser Pheromon-Antagonisten-Effekt wurde später in Feldtests bestätigt, bei denen das Einfangen von C. gnidiella-Männchen in einem dosisabhängigen Muster abnahm. Der Pheromon-Antagonismus wurde auch für B. mandarina, einen Vorfahren von B. mori (Daimon et al., 2012)., Ihre Ergebnisse bestätigen Bombykol als Sexualpheromon und Bombykal und Bombykylacetat als Antagonisten, die die Anziehungskraft von Männern im Feld auf das Sexualpheromon Bombykol stark hemmten. In jüngerer Zeit wurden Hinweise auf einen Pheromonantagonismus für die Schnauzenmotte Herpetogramma Polyginale berichtet. Wenn (Z)-13-hexadezicenol zu seinem Sexualpheromon hinzugefügt wurde, (Z)-13-Hexadezicenylacetat, verringerte sich signifikant die Anzahl der im Feld gefangenen Männchen (Yan et al., 2015)., Der Pheromon-Antagonismus scheint auf den Unterschieden in der chemischen Funktionsgruppe wie Alkoholen, Aldehyden und Estern in Abhängigkeit von der Insektenart zu beruhen. Nichtsdestotrotz ist die antagonistische Wirkung dieser Strukturanaloga möglicherweise nicht auf die ORco-Hemmung zurückzuführen, sondern auf die Spezifität von ORx gegenüber Antagonisten. Eine kürzlich durchgeführte Studie legt nahe, dass die OR16 von Helicoverpa armigera in der Lage ist, den Pheromon-Antagonisten (Z)-11-hexadezicenol (Chang H. et al., 2017). Die Autoren unterstützten die spezifische Rolle von OR16, wenn man bedenkt, dass H., armigera-Weibchen emittieren die Antagonistenverbindung zusammen mit ihrem Sexualpheromon ((Z)-11-hexadezicenal und (Z)-9-hexadezicenal) als Strategie, um eine nicht optimale Paarung mit unreifen Männchen zu vermeiden. Herausragend, wenn die OR16 wurde durch die Genom-Editing-Technik CRISPR niedergeschlagen / Cas9 und H. armigera Männchen wurden durch Elektrophysiologie und Verhaltenstests getestet, keine EAG-Reaktion wurde aufgezeichnet und Männer versuchten, mit unreifen Frauen zu paaren.
Geruchsrezeptoren vs., Bindungsproteine: Vor-und Nachteile für die Schädlingsbekämpfung von Insekten
Bei OBPs stellen die Ligandenspezifität und-mechanismen von OBPs kontroverse Aspekte dar, die stark von den Methoden zur Messung der Ligandenaffinität abhängen. Zum Beispiel wurde berichtet, dass PBPs, wie die von den Motten‘ P. xylostella und Eogystiapferhaecolus, sowohl Geschlechtspheromonkomponenten als auch Analoga binden können (Sun et al., 2013a; Hu et al., 2018). Dies deutet darauf hin, dass Downstream-Player wie ORs die Spezifität und Empfindlichkeit des Geruchsmittelempfangs verbessern könnten., Jüngste Beweise belegen, dass die Koexpression von PBPs und PRs die Empfindlichkeit gegenüber Pheromonen erhöhen kann. Beispielsweise wurden mehrere Kombinationen aus PR1-4 und PBP1-4 verwendet, um ihre Reaktion auf Sexualpheromonkomponenten der Motten-Chilo-Suppressalis zu testen (Chang et al., 2015). Die Autoren fanden eine signifikante Erhöhung der Empfindlichkeit der Reaktion in Richtung (Z)-11-hexadecenal, wenn PR4 und PR6 wurden co-exprimiert mit PBP4., Obwohl die Wechselwirkung dieser Proteine ein neues Forschungsniveau als Schädlingsbekämpfungsziele erreichen könnte, beleuchtet die unterschiedliche Paarung von PRs und PBPs die Komplexität des Geruchsystems bei Insekten, was den Ansatz zu einer schwierigen Aufgabe für eine große Anzahl von Verbindungen und Proteinen macht testen. Trotz der oben genannten, insekten OBPs sind von kleiner molekularer größe mit einfacher produktion von rekombinanten proteinen, die macht sie lieblings ziele für strukturelle studien und schnelle bindung screening., Zum Beispiel könnte das Ligandenscreening mit OBPs die Identifizierung chemischer Eigenschaften für eine bessere Bindung ermöglichen, wie Kettenlänge, Molekularvolumen, funktionelle Gruppen und Bindungsunsättigung. Diese, kombiniert mit neuen Proteinstrukturvorhersagemethoden, wie sie bei der Gestaltung von Arzneimitteln und Antikörpern verwendet werden, wie Homologiemodellierung, Dynamiksimulationen und molekulares Andocken, könnten Insektenobps gegenüber ORs als Ziele für die Entwicklung von Kontrollmitteln in der Schädlingsbekämpfung in eine bevorzugte Position bringen.
Insektenstiche scheinen spezifischer auf Geruchsstoffe als OBPs abgestimmt zu sein., Die höhere Spezifität von ORs und die Chance auf Aktivierung / Hemmung spezifischer Rezeptoren für ein bestimmtes Verhalten machen diese Proteine zu attraktiven Zielen, um Schädlingsverhalten zu manipulieren. Die Durchführbarkeit der Hemmung von entweder ORx / ORco-Komplex oder ORco durch Antagonisten umfasst eine vielversprechende Strategie, um insektenspezifisches Verhalten wie die Paarung über Sexualpheromonrezeptoren zu stören. Der Mangel an strukturellen Informationen ist jedoch der Engpass bei der Verwendung von Insektenors als Ziele für die Vorhersage semiochemischer Aktivitäten., Die Tabellen 1, 2 fassen die Anzahl der OBPs und ORs zusammen, die in Insektenarten durch Transkriptom (d. H. RNA-seq) und Genomsequenzierung identifiziert wurden. Die meisten der bisher untersuchten Insekten haben laut Genomstudien mindestens zweimal ORs als OBPs. Darüber hinaus gibt es eine umfangreiche Expansion von ORs in sozialen Insekten aus der hymenopteren Ordnung wie der Honigbiene A. mellifera mit 170 ORs (Weinstock et al., 2006)und die Ameisen Solenopsis invicta und Cerapachys biroi mit 400 bzw., 2011; Oxley et al., 2014)., In ähnlicher Weise zeigt sich die OR-Expansion auch bei einigen landwirtschaftlichen Schädlingen, wie dem Rotmehlkäfer T. castaneum mit 265 ORs im Vergleich zu 47 OBPs (Richards et al., 2008). Dies macht eine anspruchsvolle Aufgabe für das Ziel ODER die Auswahl zusammen mit der Schwierigkeit für die funktionelle Expression von Transmembranproteinen wie ORs, um eine große Anzahl von Liganden zu screenen. Eine Annäherung wichtiger Eigenschaften in OBPs und ORs ist in Tabelle 3 zusammengefasst.
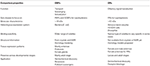
Tabelle 3., Ungefährer Vergleich von Insektenors und OBPs nach Eigenschaften.
Weitere Perspektiven
Die funktionelle Charakterisierung von Insektenors sowie ihre nachgewiesene Rolle bei der Insektengeruch beleuchten die Empfindlichkeit und Spezifität dieser insektenspezifischen Proteine. Diese Fortschritte werden ihre Machbarkeit als Schädlingsbekämpfungsziele durch das Verständnis molekularer Erkennungsmechanismen und kombinatorischer Wechselwirkungen mit OBPs weiter verbessern., Andererseits wird der derzeitige massive Aufwand bei der Identifizierung und verbindlichen Charakterisierung von OBPs in mehreren landwirtschaftlich wichtigen Insektenarten anhalten und mehr Informationen über ihre Funktionen in der Insektenphysiologie liefern. Daher schlägt diese Überprüfung als Hauptvorteil für OBPs gegenüber ORs die Verfügbarkeit von 3D-Kristall-und NMR-Strukturen vor, die mit nachgelagerten Ansätzen wie der Homologiemodellierung (falls erforderlich), dem molekularen Andocken und der Molekulardynamik die Suche nach bioaktiven Chemikalien verfeinern würden., Diese letzte in Ergänzung zur Liganden-Affinitätsmessung wird die Untersuchung von Insektenobps beschleunigen, die als Ziele für die semiochemische Entdeckung und die Werkzeuge zur Gestaltung von Superliganden im Schädlingsbekämpfungsmanagement überdacht werden sollen.
Das Auftreten und die Entwicklung von Insektizidresistenz bei Insektenschädlingen haben zu intensiver Erforschung des Insektengeruchs und der Mechanismen der neuronalen Verarbeitung geführt. Es ist bekannt, dass eine Reihe von Rezeptoren und Enzymen im INSEKTENZNS die Ziele für die Entwicklung von Insektizidresistenzen sind (Abbildung 1)., Es wurde gezeigt, dass Acetylcholinesterase (AChE) in löslicher Form die Resistenz gegen Organo-Phosphor-und Carbamat-Insektizide bietet, die als Bioscavengers wirken (Lee et al., 2015). In ähnlicher Weise wurden bei der Blattlaus M. persicae mehrere Insektizidresistenzmechanismen nachgewiesen, die Carboxylesterasen, Natriumkanäle, γ-Aminobuttersäure (GABA) und nAChR (Bass et al., 2014)., Als wichtige Komponenten im Nervensystem der Insektenperipherie und Hauptakteure im Insektenverhalten stellen sowohl Insektenschutzmittel als auch ORs alternative Ziele für die Identifizierung von Verbindungen mit semiochemischer Aktivität (oder Agonisteneffekt) und Werkzeugen zur Entwicklung starker Antagonisten dar, um die gewünschten Verhaltensreaktionen von Insektenschädlingen zu verbessern und den Einsatz von Insektiziden und anschließender Resistenz zu reduzieren.
Autorenbeiträge
HV schrieb Abschnitte über Struktur und ORs von OBP, entwickelte Tabellen und Abbildungen., J-JZ konzipierte die Idee für den Übersichtsartikel, schrieb den Hauptteil, wie Einführung, OBPs-Funktion und-Struktur sowie OR-OBP-Vergleich.
Interessenkonflikterklärung
Die Autoren erklären, dass die Untersuchung ohne kommerzielle oder finanzielle Beziehungen durchgeführt wurde, die als potenzieller Interessenkonflikt ausgelegt werden könnten.
Danksagungen
Die Autoren danken FONDECYT 3170433., J-JZ ist dankbar für die finanzielle Unterstützung von Northeast Normal University und Jilin University, China für seinen sabbatical Urlaub in China zu studieren. Rothamsted Research erhält ein Stipendium mit zusätzlichen Mitteln aus Biotechnologie und Biowissenschaften (BBSRC), UK.
– Gonzalez, F., Witzgall, P., and Walker, W. B. (2017). Antennentranskriptome von drei tortricid Motten zeigen mutmaßlich konservierte chemosensorische Rezeptoren für soziale und Lebensraum olfaktorische Hinweise. Sci. Rep. 7, 1-12. doi: 10.,1038/srep41829
PubMed Abstract | CrossRef Full Text | Google Scholar
Kaissling, K. E. (2013). Die Kinetik der Geruchsreaktionen könnte weitgehend von der Geruchsrezeptor-Wechselwirkung und der für Flussmitteldetektoren postulierten Geruchsdeaktivierung abhängen. J. Comp. Physiol. Neuroethol. Sensorisches neuronales Verhalten. Physiol. 199, 879–196. doi: 10.1007 / s00359-013-0812-z
PubMed Abstract / CrossRef Volltext / Google Scholar