chociaż lotne związki o aktywności agonistycznej zostały przebadane przeciwko ORs, specyficznej substancji chemicznej o silnym działaniu agonistycznym na komary, 2-(4-etylo-5-(pirydyn-3-ylo)-4h-1,2,4-triazol-3-ylotio)-N-(4-etylofenylo)acetamid (VUAA1), otworzył pole badań (Jones et al., 2011; Taylor et al., 2012). Późniejsze badania Taylor et al., (2012) dostarczył dowodów na chemikalia pochodne VUAA1, takie jak VUAA4, zdolne do zwiększenia jego działania agonistycznego 10-krotnie na ORco z A. gambiae, H. virescens i Harpegnathos saltator. Co ciekawe, autorzy donosili, że jakakolwiek zmiana podstawników amidowych spowoduje całkowitą utratę aktywności agonistycznej. Daje to pomocne spostrzeżenia na temat wymagań strukturalnych agonistów i relacji struktura-aktywność między analogami VUAA i RNO. Wreszcie, pomimo zwiększonej aktywności agonistycznej substancji chemicznych vuaa, jej stosunkowo wysoka masa cząsteczkowa (367,47 g mol−1 dla VUAA1) vs., lotne agonisty, takie jak bombykol (238,42 g mol−1), sprawia, że bezpośrednie lotne dostarczenie VUAA jest niemożliwe. Mając to na uwadze, poszukiwanie mniejszych analogów strukturalnych stanowi interesujący przedmiot badań.
antagonizm wobec podjednostki ORco
wraz z badaniem analogów związanych z VUAA, które mogą działać jako silne agonisty, pojawiła się również blokada ORco przez antagonistów, aby kierować semiochemikaliami i projektowaniem pestycydów., Tak więc, strukturalny analog VUAA1, VU0183254 (2-(4-etylo-5-furan-2-ylo-4H-triazol-3-ylsulfanylo)-1-fenotiazyn-10-ylo-etanonu), zgłoszono hamowanie odpowiedzi ORco, działając jako allosteryczny modulator w A. gambiae i zakłócając rozpoznawanie agonistów takich jak eugenol przez kompleks OR65/ORco (Jones et al., 2012). Inne analogi strukturalne VUAA były również zgłaszane jako antagoniści. Przykładem są N -, 2-podstawione związki triazolotioacetamidu OLC3 i OLC12, które zakłócają reakcję ORco w podobny sposób u C. quinquefasciatus, A. gambiae, D. melanogaster I O., nubilalis, sugerując zachowane miejsce wiązania w ORco (Chen i Luetje, 2012). Biorąc pod uwagę hamowanie Orco jako obiecującą strategię zakłócania zachowań owadów, wydaje się, że późniejsze wysiłki powinny zmierzać do związków o mniejszej masie cząsteczkowej niż antagoniści pochodni VUAA. Na przykład stwierdzono, że OX1a (232 g mol−1), tryptamina (160,22 g mol−1) i cynamonian izopropylu (190,24 g mol−1) mają działanie antagonistyczne na ORco (Chen and Luetje, 2013, 2014; Tsitoura et al., 2015) o około połowie lub mniejszej masie cząsteczkowej niż VUAA1., Niemniej jednak należy dokładnie zbadać przyszłe stosowanie tych antagonistów, ponieważ zablokowanie konserwowanego ORco może wpływać nie tylko na szkodliwe owady, ale także na pożyteczne.
poza działaniem antagonistycznym badanym in vitro, dowody na poziomie behawioralnym wspierają ideę, że strukturalne analogi feromonów mogą funkcjonować jako antagoniści. Na przykład Sellanes et al., (2010) poinformował o hamowaniu reakcji seksualnej w cryptoblabes gnidiella, gdy analogi strukturalne, mrówczan (Z)-9-tetradecenylu i mrówczan (Z)-11-heksadecenylu, dodano do syntetycznych feromonów płciowych, (Z)-11-heksadecenal i (Z)-13-oktadecenal, w badaniach w tunelu aerodynamicznym. To działanie antagonistyczne feromonów zostało później potwierdzone w testach terenowych, gdzie pułapkowanie samców C. gnidiella zmniejszyło się w zależności od dawki. Antagonizm feromonów odnotowano również dla B. mandarina, przodka B. mori (Daimon et al., 2012)., Ich odkrycia potwierdzają bombykol jako feromon płciowy, a bombykal i octan bombykylu jako antagoniści, które silnie hamowały przyciąganie samców w polu do feromonu płciowego bombykol. Niedawno, dowody antagonizmu feromonów donoszono o pysk ćmy Herpetogramma submarginale. Kiedy (Z)-13-heksadecenol dodano do jego feromonu płciowego, (z) – 13-octan heksadecenylu, znacznie zmniejszyła się liczba mężczyzn schwytanych w polu (Yan et al., 2015)., Antagonizm feromonów wydaje się oparty na różnicach w chemicznych grupach funkcyjnych, takich jak alkohole, aldehydy i estry w zależności od gatunku owadów. Niemniej jednak działanie antagonistyczne tych analogów strukturalnych może być spowodowane nie hamowaniem ORco, ale swoistością ORx dla antagonistów. Ostatnie badania sugerują, że OR16 Helicoverpa armigera jest w stanie konkretnie rozpoznać antagonistę feromonów, (Z)-11-heksadecenol (Chang H. et al., 2017). Autorzy poparli szczególną rolę OR16, uznając, że H., samice armigera emitują związek antagonistyczny wraz z feromonem płciowym ((z)-11-heksadecenal i (Z) – 9-heksadecenal) jako strategię unikania nieoptymalnego kojarzenia z niedojrzałymi samcami. Co ciekawe, kiedy OR16 został powalony przez technikę edycji genomu CRISPR / Cas9 i H. armigera samce były testowane przez testy elektrofizjologiczne i behawioralne, nie zarejestrowano żadnej odpowiedzi EAG, a mężczyźni próbowali kojarzyć się z niedojrzałymi kobietami.
Receptory zapachowe a, Binding Proteins: plusy i minusy dla Insect Pest Management
w przypadku OBPs specyficzność ligandowa i mechanizmy OBPs reprezentują kontrowersyjne aspekty, co wydaje się silnie zależne od metod stosowanych do pomiaru powinowactwa ligandowego. Na przykład, donoszono, że PBP, takie jak te z ćmy ” P. xylostella i eogystia hippophaecolus, może wiązać zarówno feromony płciowe składniki i analogi (Sun et al., 2013a; Hu i in., 2018). Sugeruje to, że odtwarzacze niższego szczebla, takie jak RNO, mogą zwiększyć swoistość i wrażliwość odbioru odorantów., Najnowsze dowody potwierdzają, że współekspresja PBPs i PRs może zwiększyć wrażliwość na feromony. Na przykład, wiele kombinacji z PR1-4 i PBP1-4 zostały wykorzystane do testowania ich odpowiedzi na składniki feromonów płciowych ćmy Chilo suppressalis (Chang et al., 2015). Autorzy stwierdzili znaczny wzrost czułości odpowiedzi na (z)-11-heksadecenal, gdy PR4 i PR6 były jednocześnie wyrażane z PBP4., Chociaż interakcja tych białek może powstać nowy poziom badań jako celów zwalczania szkodników, różne parowanie PRs i PBP rzuca światło na złożoność systemu węchowego u owadów, czyniąc podejście trudnym zadaniem dla dużego zestawu związków i białek do przetestowania. Pomimo powyższego, owad OBP są małych rozmiarów molekularnych z łatwą produkcją rekombinowanych białek, co czyni je ulubionymi celami Badań Strukturalnych i szybkiego przesiewania wiązania., Na przykład, ligand przesiewanie z OBPs może pozwolić na identyfikację właściwości chemicznych dla lepszego wiązania, takich jak długość łańcucha, objętość cząsteczkowa, funkcjonalne grupy i wiązania nienasycenie. Te, w połączeniu z nowymi metodami przewidywania struktury białek, stosowanymi w projektowaniu leków medycznych i przeciwciał, takimi jak modelowanie homologiczne, symulacje dynamiki i dokowanie molekularne, mogą umieścić OBP owadów w ulubionej pozycji nad ORs jako cele dla rozwoju środków kontrolnych w zarządzaniu szkodnikami.
insekty wydają się bardziej dostrojone do substancji zapachowych niż OBP., Wyższa specyficzność wykazywana przez Or i szansa na aktywację / zahamowanie określonych receptorów dla danego zachowania sprawiają, że białka te są atrakcyjnymi celami do manipulowania zachowaniami szkodników. Możliwość hamowania kompleksu ORx/ORco lub Orco przez antagonistów obejmuje obiecującą strategię zakłócania specyficznych zachowań owadów, takich jak kojarzenie się przez receptory feromonów płciowych. Jednak brak informacji strukturalnych jest wąskim gardłem w wykorzystywaniu insektów jako celów do przewidywania aktywności semiochemicznej., Tabele 1, 2 podsumowują liczbę OBP i RNO, które zostały zidentyfikowane u gatunków owadów za pomocą transkryptomu (tj. RNA-seq) i sekwencjonowania genomu. Większość badanych do tej pory owadów ma co najmniej dwa ORs niż OBPs zgodnie z badaniami genomu. Ponadto występuje rozległa ekspansja Or w owadach społecznych z rzędu Hymenoptera, takich jak pszczoła miodna A. mellifera z 170 or (Weinstock et al., 2006), oraz Mrówki Solenopsis invicta i Cerapachys biroi z 400 i 506 or, odpowiednio (Wurm et al., 2011; Oxley et al., 2014)., Podobnie ekspansja OR jest również widoczna w niektórych szkodnikach rolniczych, takich jak chrząszcz z czerwonej mąki T. castaneum z 265 RNO w porównaniu do 47 OBPs (Richards et al., 2008). To sprawia, że wymagające zadanie dla celu lub selekcji wraz z trudnością dla funkcjonalnej ekspresji białek transmembranowych, takich jak ORs, w celu ekranowania dużej liczby ligandów. Przybliżenie ważnych właściwości zarówno OBPs, jak i RNO podsumowano w tabeli 3.
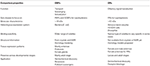
Tabela 3., Przybliżone porównanie Or i OBP owadów według właściwości.
dalsze perspektywy
funkcjonalna charakterystyka RNO owadów oraz ich udowodniona rola w węchu owadów rzuciły światło na czułość i swoistość tych białek specyficznych dla owadów. Postępy te jeszcze bardziej zwiększą ich wykonalność jako celów zwalczania szkodników poprzez zrozumienie mechanizmów rozpoznawania molekularnego i kombinacyjnych interakcji z OBPs., Z drugiej strony, obecne masowe wysiłki w identyfikacji i wiążącej charakterystyce OBPs w kilku ważnych dla rolnictwa gatunkach owadów będą kontynuowane i dostarczą więcej informacji na temat ich funkcji w fizjologii owadów. Tak więc w niniejszym przeglądzie proponuje się jako główną zaletę dla OBPs nad RNO, dostępność struktur krystalicznych 3D i NMR, które przy podejściach niższego szczebla, takich jak modelowanie homologiczne (w razie potrzeby), dokowanie molekularne i dynamika molekularna, udoskonaliłyby poszukiwania bioaktywnych substancji chemicznych., To ostatnie uzupełnienie pomiaru powinowactwa ligandowego przyspieszy badania OBP owadów, które zostaną ponownie rozważone jako cele do odkrycia semiochemicznego i narzędzia do projektowania super-ligandów w zarządzaniu szkodnikami.
pojawienie się i rozwój odporności owadobójczej u szkodników owadów doprowadziło do intensywnych badań nad węchem owadów i mechanizmami, które są zaangażowane w przetwarzanie nerwowe. Ustalono, że wiele receptorów i enzymów w OUN owadów jest celami rozwoju odporności na insektycydy (ryc. 1)., Wykazano, że acetylocholinoesteraza (AChE) w formie rozpuszczalnej zapewnia odporność na insektycydy organofosforowe i karbaminianowe, działając jako bioscavengers (Lee et al., 2015). Podobnie wykazano wiele mechanizmów odporności na insektycydy u mszycy M. persicae, obejmujących karboksyloesterazy, kanały sodowe, kwas γ-aminomasłowy (GABA) i Nachr (Bass et al., 2014)., Jako ważne składniki w obwodowym układzie nerwowym owadów i kluczowych graczy w zachowaniach owadów, zarówno owad OBPs i RNO stanowią alternatywne cele do identyfikacji związków o aktywności semiochemicznej (lub efekt agonistyczny) i narzędzia do projektowania silnych antagonistów w celu zwiększenia pożądanych reakcji behawioralnych szkodników owadów i zmniejszenia stosowania środków owadobójczych i późniejszej odporności.
wkład autora
HV napisał sekcje o strukturze OBP i ORs, opracował tabele i ryciny., J-JZ wpadł na pomysł artykułu przeglądowego, napisał sekcję główną, taką jak wprowadzenie, funkcja i struktura OBPs oraz porównanie OR-OBP.
Oświadczenie o konflikcie interesów
autorzy oświadczają, że badanie zostało przeprowadzone w przypadku braku jakichkolwiek relacji handlowych lub finansowych, które mogłyby być interpretowane jako potencjalny konflikt interesów.
podziękowania
autorzy pragną podziękować FONDECYT 3170433., J-JZ jest wdzięczny za wsparcie finansowe Northeast Normal University i Jilin University w Chinach na jego urlop naukowy na studia w Chinach. Rothamsted Research otrzymuje grant z dodanym finansowaniem z biotechnologii i Nauk Biologicznych (BBSRC), Wielka Brytania.
Transcriptomes Antennal trzy tortricid ćmy ujawniać przypuszczalny konserwujący chemosensory receptory dla społecznego i siedliska węchowych sygnałów. Sci. REP. 7, 1-12. doi: 10.,1038 / srep41829
PubMed Abstract / CrossRef Full Text / Google Scholar
Kinetyka odpowiedzi węchowych może w dużej mierze zależeć od interakcji odorant-receptor i dezaktywacji zapachowej postulowanej dla detektorów strumienia. J. Comp. Physiol. Neuroethol. Zmysłowe Zachowania Nerwowe. Physiol. 199, 879–196. podoba mi się! do obserwowanych nr: 60359-013-0812-z
PubMed Abstract | CrossRef Full Text | Google Scholar