Cíle Vzdělávání
- Porovnat získaná operons a repressible operons
- Popsat, proč nařízení operons je důležité
Každý nucleated buněk v mnohobuněčný organismus obsahuje kopie stejnou DNA. Podobně, všechny buňky ve dvou čistých bakteriálních kultur inokulovaných ze stejné výchozí kolonie obsahují stejnou DNA, s výjimkou změn, které vznikají spontánní mutace., Pokud má každá buňka v mnohobuněčném organismu stejnou DNA, jak je možné, že buňky v různých částech těla organismu vykazují různé vlastnosti? Podobně, jak je možné, že stejné bakteriální buňky ve dvou čistých kulturách vystavených různým podmínkám prostředí mohou vykazovat různé fenotypy? V obou případech se každá geneticky identická buňka nezapne ani nevyjadřuje stejnou sadu genů. Vyjadřuje se pouze podmnožina proteinů v buňce v daném čase.,
Genomické DNA obsahuje strukturní geny, které kódují produkty, které slouží jako buněčné struktury nebo enzymy a regulační geny, které kódují produkty, které regulují genovou expresi. Exprese genu je vysoce regulovaný proces. Zatímco regulace genové exprese v mnohobuněčných organismech umožňuje buněčnou diferenciaci, v jednobuněčných organismech, jako jsou prokaryoty, primárně zajišťuje, že zdroje buňky nejsou zbytečné, takže proteiny, které buňka v té době nepotřebuje.,
objasnění mechanismů kontrolujících genovou expresi je důležité pro pochopení lidského zdraví. Poruchy v tomto procesu u lidí vedou k rozvoji rakoviny a dalších onemocnění. Pochopení interakce mezi genovou expresí patogenu a interakcí jeho lidského hostitele je důležité pro pochopení konkrétní infekční nemoci. Genová regulace zahrnuje komplexní síť interakcí uvnitř dané buňky mezi signály z buněčného prostředí, signalizační molekuly v buňce a DNA buňky., Tyto interakce vedou k expresi některých genů a potlačení ostatních, v závislosti na okolnostech.
Prokaryot a eukaryot sdílejí některé podobnosti v jejich mechanismy, které regulují genovou expresi; nicméně, genové exprese u eukaryot je mnohem složitější, protože časové a prostorové oddělení mezi procesy transkripce a translace., Takže, i když většina regulaci genové exprese se vyskytuje přes transkripční kontrolu v prokaryot, regulace genové exprese u eukaryot dochází na transkripční úrovni a post-transkripčně (po primární přepis byl učiněn).
Prokaryotické Genové Nařízení
U bakterií a archaea, strukturální proteiny s související funkce jsou obvykle kódovány v genomu v bloku nazývá operon a jsou přepisovány společně pod kontrolou jednoho promotoru, což má za následek vznik polycistronic přepis (Obrázek 1)., Tímto způsobem, regulace transkripce všech strukturálních genů kódujících enzymy, které katalyzují mnoho kroků v jedné biochemické dráhy může být ovládán současně, protože buď budou všechny být potřebné ve stejné době, nebo žádná nebude potřeba. Například, v E. coli, všechny strukturní geny, které kódují enzymy potřebné k využití laktózy jako zdroje energie ležet vedle sebe v laktóza (nebo lac) operon pod kontrolou jednoho promotoru, lac promotor., Francouzští vědci François Jacob (1920-2013) a Jacques Monod v Pasteurově Institutu byly nejprve ukázat organizaci bakteriální geny do operons, prostřednictvím svých studií na lac operon E. coli. Za tuto práci získali v roce 1965 Nobelovu cenu za fyziologii nebo medicínu. Ačkoli eukaryotické geny nejsou organizovány do operantů, prokaryotické operony jsou vynikajícími modely pro učení o regulaci genů obecně. V eukaryotách existují některé genové klastry, které fungují podobně jako operony., Mnoho principů lze aplikovat na eukaryotické systémy a přispět k našemu pochopení změn genové exprese v eukaryotách, které mohou vést k patologickým změnám, jako je rakovina.

Obrázek 1. V prokaryot, strukturální geny související funkce jsou často organizovány společně na genomu a přepsal společně pod kontrolou jednoho promotoru. Regulační oblast operonu zahrnuje jak pořadatele, tak provozovatele. Pokud se represor váže na operátora, nebudou strukturní geny přepsány., Alternativně se aktivátory mohou vázat na regulační oblast, což zvyšuje transkripci.
každý operon obsahuje sekvence DNA, které ovlivňují jeho vlastní transkripci; ty se nacházejí v oblasti zvané regulační oblast. Regulační oblast zahrnuje promotor a oblast obklopující promotor, na kterou se mohou vázat transkripční faktory, proteiny kódované regulačními geny. Transkripční faktory ovlivňují vazbu RNA polymerázy na promotor a umožňují jeho progresi přepisovat strukturální geny., A repressor je transkripční faktor, který potlačuje transkripci genu v reakci na externí stimul vazbou na sekvenci DNA v regulační oblasti zvané provozovatel, který se nachází mezi RNA polymeráza vazebné místo promotoru a transkripční start místě první strukturální gen. Represorová vazba fyzicky blokuje RNA polymerázu z přepisu strukturních genů., Naopak aktivátor je transkripční faktor, který zvyšuje transkripci genu v reakci na vnější stimul tím, že usnadňuje vazbu RNA polymerázy na promotor. Induktor, třetí typ regulační molekuly, je malá molekula, která buď aktivuje nebo potlačuje transkripci interakcí s represorem nebo aktivátorem.
u prokaryot existují příklady opronů, jejichž genové produkty jsou vyžadovány spíše důsledně a jejichž exprese je proto neregulovaná., Tyto operony jsou konstitutivně vyjádřeny, což znamená, že jsou transkribovány a přeloženy nepřetržitě, aby buňce poskytly konstantní mezilehlé hladiny proteinových produktů. Tyto geny kódují enzymy, které se podílejí na úklidových funkcích potřebných pro buněčnou údržbu, včetně replikace DNA, opravy a exprese, jakož i enzymů podílejících se na metabolismu jádra. Naproti tomu existují další prokaryotické operony, které jsou vyjádřeny pouze v případě potřeby a jsou regulovány represory, aktivátory a induktory.,
Přemýšlejte o tom
- jaké jsou části sekvence DNA operonu?
- jaké typy regulačních molekul existují?
Nařízení o Represi
Prokaryotické operons jsou běžně kontrolována vazba repressors obsluhy regionů, čímž se zabrání přepisu strukturních genů. Tyto operony jsou klasifikovány buď jako represivní operony, nebo jako indukovatelné operony. Represivní operony, jako je tryptofan (trp) operon, obvykle obsahují geny kódující enzymy potřebné pro biosyntetickou cestu., Dokud bude produkt dráhy, jako je tryptofan, nadále vyžadován buňkou, bude i nadále vyjádřen represivní operon. Nicméně, když produkt biosyntetické dráhy začne hromadit v buňce, odstraní potřebu pro buňky, aby i nadále, aby se více, výraz operon je potlačena. Naopak indukovatelné operony, jako je Lac operon E. coli, často obsahují geny kódující enzymy v cestě zapojené do metabolismu specifického substrátu, jako je laktóza., Tyto enzymy jsou vyžadovány pouze tehdy, když je tento substrát k dispozici, takže exprese operonu je obvykle indukována pouze za přítomnosti substrátu.
trp Operon: Repressible Operon
E. coli může syntetizovat tryptofanu pomocí enzymů, které jsou kódovány pěti strukturních genů se nachází vedle sebe v trp operon (Obrázek 2). Když je environmentální tryptofan nízký, operon se zapne. To znamená, že transkripce je iniciována, geny jsou exprimovány a tryptofan je syntetizován., Pokud je však tryptofan přítomen v prostředí, je operon trp vypnutý. Transkripce se nevyskytuje a tryptofan není syntetizován.
když tryptofan není přítomen v buňce, represor sám o sobě se neváže na operátora; proto je operon aktivní a tryptofan je syntetizován. Nicméně, když se tryptofan hromadí v buňce, dvě molekuly tryptofanu se vážou na molekulu TRP represoru, která mění svůj tvar, což jí umožňuje vázat se na operátora trp., Tato vazba aktivní formy TRP represoru na operátora blokuje RNA polymerázu z přepisu strukturních genů a zastavuje expresi operonu. Skutečný produkt biosyntetické dráhy řízené operonem tedy reguluje expresi operonu.

Obrázek 2. Pět strukturních genů potřebných k syntéze tryptofanu v e. coli se nachází vedle sebe v operonu trp. Když tryptofan chybí, represorový protein se neváže na operátora a geny jsou transkribovány., Když je tryptofan hojný, tryptofan váže represorový protein v operátorské sekvenci. To fyzicky blokuje RNA polymerázu z přepisu genů biosyntézy tryptofanu.
podívejte se na toto video a dozvíte se více o operonu trp.
lac Operon: Indukovatelných Operon
lac operon je příklad indukovatelných operon, který je také předmětem k aktivaci v nepřítomnosti glukózy (Obrázek 3)., Lac operon kóduje tři strukturální geny nezbytné pro získání a zpracování disacharidové laktózy z prostředí a rozkládá ji na jednoduché cukry glukózu a galaktózu. Aby byl Lac operon vyjádřen, musí být přítomna laktóza. To dává smysl pro buňku, protože by bylo energeticky zbytečné vytvářet enzymy pro zpracování laktózy, pokud by laktóza nebyla k dispozici.
V nepřítomnosti laktózy, lac repressor je povinen provozovatel regionu lac operon, fyzicky brání RNA polymerázy od přepisu strukturních genů., Pokud je však přítomna laktóza, laktóza uvnitř buňky se převede na alolaktózu. Allolactose slouží jako induktor molekuly, vazba na repressor a mění svůj tvar tak, že je již schopen vázat se na operátor DNA. Odstranění repressor v přítomnosti laktózy umožňuje RNA polymeráza se pohybovat přes operátora regionu a zahájit transkripci lac strukturální geny.
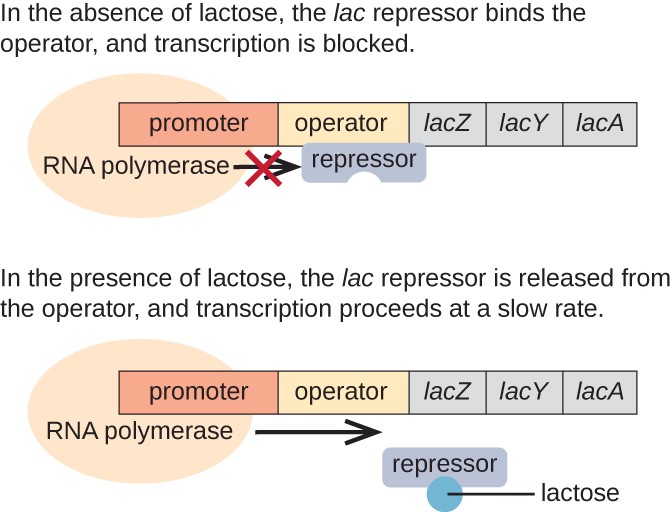
obrázek 3. Tři strukturální geny, které jsou potřebné k degradaci laktózy v e. coli, se nacházejí vedle sebe v Lac operonu., Když laktóza chybí, represorový protein se váže na operátora a fyzicky blokuje RNA polymerázu z přepisu strukturálních genů lac. Když se laktóza je k dispozici, laktóza molekula váže repressor protein, brání repressor z vazby na provozovatele sekvence a geny jsou přepisovány.
lac Operon: Aktivace Catabolite Activator Protein

Obrázek 4. Při pěstování v přítomnosti dvou substrátů používá E. coli preferovaný substrát (v tomto případě glukózu), dokud není vyčerpán., Poté jsou vyjádřeny enzymy potřebné pro metabolismus druhého substrátu a růst pokračuje, i když pomaleji.
bakterie mají obvykle schopnost používat různé substráty jako zdroje uhlíku. Protože je však glukóza obvykle výhodnější než jiné substráty, mají bakterie mechanismy, které zajišťují, že alternativní substráty se používají pouze tehdy, když byla glukóza vyčerpána. Kromě toho mají bakterie mechanismy, které zajišťují, že geny kódující enzymy pro použití alternativních substrátů jsou vyjádřeny pouze tehdy, je-li k dispozici alternativní substrát., V roce 1940, Jacques Monod byl první, ukazují preference pro určité substráty nad ostatními, přes jeho studia E. coli růst při kultivaci v přítomnosti dvou různých substrátů současně. Takové studie generovaly diauxické růstové křivky, jako je znázorněno na obrázku 4. Ačkoli se nejprve používá preferovaná substrátová glukóza, E. coli rychle roste a enzymy pro metabolismus laktózy chybí. Nicméně, jakmile hladina glukózy v krvi jsou vyčerpány, tempo růstu pomalé, navozující expresi enzymů potřebných pro metabolismus druhého substrátu, laktóza., Všimněte si, jak je rychlost růstu laktózy pomalejší, jak je naznačeno nižší strmostí růstové křivky.
schopnost přejít z použití glukózy na jiný substrát, jako je laktóza, je důsledkem aktivity enzymu nazývaného enzym IIA (EIIA). Když hladiny glukózy klesají, buňky produkují méně ATP z katabolismu (viz katabolismus sacharidů) a EIIA se stává fosforylovanou., Fosforylované EIIA aktivuje adenylyl cyklázy, enzymu, který převádí některé zbývající ATP na cyklické AMP (cAMP), cyklický derivát AMP a důležité signální molekuly podílející se na glukózu a energetický metabolismus u E. coli. Výsledkem je, že úroveň tábora začíná stoupat v buňce (obrázek 5).

obrázek 5. Když ATP pokles hladiny v důsledku vyčerpání glukózy, některé zbývající ATP se přemění na táboře adenylyl cyklázy. Zvýšené hladiny táborů tak signalizují vyčerpání glukózy.,
Lac operon také hraje roli v tomto přechodu od použití glukózy k použití laktózy. Když je glukóza vzácná, akumulační tábor způsobený zvýšenou aktivitou adenylyl cyklázy se váže na protein aktivátoru katabolitu (CAP), známý také jako protein cAMP receptor (CRP). Komplex se váže na promotorovou oblast operonu lac (obrázek 6). V regulačních oblastech těchto operonů je vazebné místo uzávěru umístěno před vazebným místem RNA polymerázy v promotoru., Vazba komplexu CAP-cAMP na toto místo zvyšuje vazebnou schopnost RNA polymerázy do promotorové oblasti iniciovat transkripci strukturních genů. V případě Lac operonu tedy musí být přítomna transkripce laktózy (odstranění Lac represorového proteinu) a hladiny glukózy musí být vyčerpány (což umožňuje vazbu aktivačního proteinu). Když jsou hladiny glukózy vysoké, dochází k katabolitové represi operonů kódujících enzymy pro metabolismus alternativních substrátů., Kvůli nízkým úrovním tábora za těchto podmínek není dostatečné množství komplexu CAP-cAMP pro aktivaci transkripce těchto operanů. Souhrn nařízení o lac operonu viz tabulka 1.

obrázek 6. a) v přítomnosti cAMP se CAP váže na promotory operonů, jako je Lac operon, které kódují geny pro enzymy pro použití alternativních substrátů. b) aby byl lac operon vyjádřen, musí být aktivován pomocí cAMP-CAP, jakož i odstranění lac represoru od provozovatele.,
| Tabulka 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
- Co ovlivňuje vazba trp operon repressor na operátora?
- jak a kdy se mění chování proteinu lac represoru?
- kromě toho, že je represivní, jak jinak je Lac operon regulován?
Globální Reakce Prokaryot
V prokaryot, tam jsou také několik vyšších úrovních genové regulace, které mají schopnost kontrolovat transkripci mnoha souvisejících operons současně v reakci na environmentální signál., Skupina operonů, které jsou současně kontrolovány, se nazývá regulon.
Alarmones
při snímání hrozícího stresu mění prokaryoti výraz široké škály opronů, aby reagovali koordinovaně. Dělají to prostřednictvím produkce alarmonů, což jsou malé intracelulární nukleotidové deriváty. Alarmony mění, které geny jsou exprimovány, a stimulují expresi specifických genů stresové reakce. Použití alarmonů ke změně genové exprese v reakci na stres se zdá být důležité u patogenních bakterií., Na setkání hostitele obranné mechanismy a další tvrdé podmínky během infekce, mnoho operons kódování virulence geny jsou upregulated v reakci na alarmone signalizace. Poznání těchto reakcí je klíčové, aby byl schopen plně porozumět procesu infekce mnoha patogenů a na vývoj terapií proti tomuto procesu.,
Alternativní sigma Faktory
Vzhledem k σ podjednotka bakteriální RNA polymerázy přiznává specifičnost, které organizace by měly být přepsány, mění σ faktor používán, je další způsob, jak pro bakterie, aby se rychle a globálně změnit to, co regulons jsou přepisována v daném čase. Faktor σ rozpoznává sekvence v bakteriálním promotoru, takže různé faktory σ rozpoznají mírně odlišné promotorové sekvence., Tímto způsobem, když buňka smysly specifické podmínky prostředí, to mohou reagovat změnou, která σ faktor vyjadřuje, ponižující staré a vyrábějí nové přepsat operons kódování genů, jejichž produkty budou užitečné v rámci nového životního stavu. Například, v sporulating bakterie rodů Bacillus a Clostridium (které zahrnují mnoho patogenů), skupina σ faktory řídí expresi mnoha genů potřebných pro sporulace v reakci na sporulace-stimulační signály.,
Přemýšlejte o tom
- jaký je název souboru operanů, které lze regulovat jako skupinu?
- jaký typ stimulu by vyvolal transkripci jiného faktoru σ?
Další Metody Regulace ve Bakterie: Útlum a Riboswitches
i když většina genové exprese je regulována na úrovni transkripce zahájení v prokaryot, tam jsou také mechanismy kontroly dokončení transkripce, stejně jako překlad souběžně., Od jejich objevu bylo prokázáno, že tyto mechanismy kontrolují dokončení transkripce a překladu mnoha prokaryotických operantů. Protože tyto mechanismy přímo spojují regulaci transkripce a překladu, jsou specifické pro prokaryoty, protože tyto procesy jsou fyzicky odděleny v eukaryotech.
Jeden takový regulační systém je útlum, přičemž sekundární stem-loop struktury, vytvořené v rámci 5′ konec mRNA je přepisována určit, zda přepis dokončit syntézu mRNA dojde a pokud je tato mRNA budou použity pro překlad., Za transkripční represe mechanismus již bylo řečeno, útlum ovládá také exprese trp operon u E. coli (Obrázek 7). Trp operon regulační oblast obsahuje vedoucí sekvenci zvané trpL mezi provozovatelem a první strukturální gen, který má čtyři úseky RNA, které mohou základní pár s sebou v různých kombinacích. Když terminátor vlásenkové formy, transkripce končí, uvolnění RNA polymerázy z mRNA., Nicméně, když antiterminator vlásenkové formy, to zabraňuje tvorbě terminátor stem-loop, tak RNA polymeráza může přepisu strukturních genů.
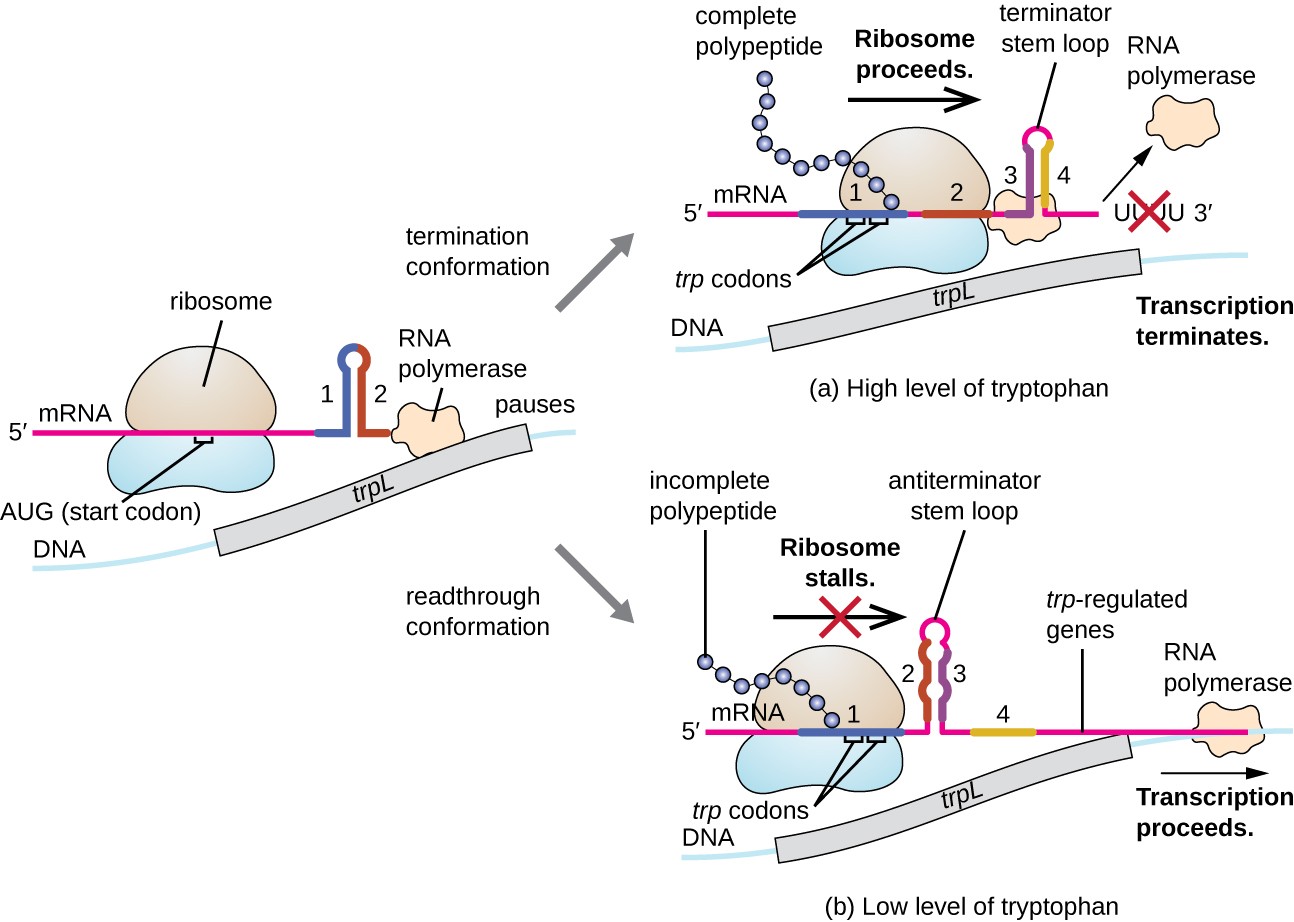
Obrázek 7. Kliknutím zobrazíte větší obrázek. Když je tryptofan hojný, pokračuje překlad peptidu krátkého vůdce kódovaného trpL, terminátorová smyčka mezi regiony 3 a 4 formy a transkripce končí., Kdy tryptofan úrovně jsou vyčerpány, překlad krátké leader peptid stánky v regionu 1, které umožňují regiony 2 a 3 tvoří antiterminator smyčky, a RNA polymeráza může přepsat strukturální geny trp operon.
související mechanismus souběžné regulaci transkripce a translace u prokaryot je použití riboswitch, malé oblasti noncoding RNA nalézt v rámci 5′ konci některé prokaryotické mRNA molekuly (Obrázek 8). Riboswitch se může vázat na malou intracelulární molekulu, aby stabilizoval určité sekundární struktury molekuly mRNA., Vazba malé molekuly určuje, která struktura kmenové smyčky se tvoří, což ovlivňuje dokončení syntézy mRNA a syntézy proteinů.

Obrázek 8. Klikněte pro větší obrázek. Riboswitches nalézt v prokaryotické mRNA molekuly se může vázat na malé intracelulární molekuly, stabilizace některých RNA struktur, které ovlivňují buď dokončení syntézy mRNA molekula sama o sobě (doleva) nebo protein vyrobený pomocí této mRNA (vpravo).,
Další Faktory, které Ovlivňují Genové Exprese u Prokaryot a Eukaryot
i když zaměření na naše diskuse o transkripční kontrolou používá prokaryotické operons jako příklady, eukaryotické transkripční ovládání je podobné v mnoha ohledech. Stejně jako u prokaryot může být eukaryotická transkripce řízena vazbou transkripčních faktorů včetně represorů a aktivátorů., Zajímavé je, že eukaryotické transkripce může být ovlivněna vazba proteinů na regiony DNA, tzv. enhancery, poměrně daleko od genu, prostřednictvím DNA opakování usnadněno mezi zesilovač a promotor (Obrázek 9). Celkově je regulace transkripce vysoce účinným způsobem kontroly genové exprese jak u prokaryot, tak u eukaryot. Kontrola genové exprese v eukaryotách v reakci na environmentální a buněčné napětí však může být provedena dalšími způsoby bez vazby transkripčních faktorů na regulační oblasti.,

obrázek 9. V eukaryotách je zesilovač sekvence DNA, která podporuje transkripci. Každý zesilovač je tvořen krátkými sekvencemi DNA nazývanými distální kontrolní prvky. Aktivátory vázané na distální kontrolní prvky interagují s mediátorovými proteiny a transkripčními faktory. Dva různé geny mohou mít stejný promotor, ale různé distální kontrolní prvky, umožňující diferenciální genovou expresi.,
DNA-Ovládání Úrovně
U eukaryot DNA molekuly nebo související histony mohou být chemicky upraveny tak, aby vliv transkripce; to se nazývá epigenetické regulace. Metylace určitých cytosin nukleotidů v DNA v reakci na faktory životního prostředí bylo prokázáno, že vliv použití těchto DNA pro transkripci, DNA metylace často koreluje se sníženou úrovní genové exprese., Navíc, v reakci na faktory životního prostředí, histon bílkovin pro balení DNA může být také chemicky upravené v několika způsoby, včetně acetylace a deacetylation, ovlivňování obalu stav DNA a tím ovlivňuje dostupnost volně navinuta DNA pro transkripci. Tyto chemické modifikace mohou být někdy udržovány prostřednictvím několika kol buněčného dělení, takže alespoň některé z těchto epigenetických změn jsou dědičné.
toto video popisuje, jak epigenetická regulace řídí genovou expresi.,
Přemýšlejte o tom
- co zastaví nebo umožňuje přepis pokračovat, když je útlum v provozu?
- co určuje stav riboswitchu?
- popsat funkci zesilovače.
- popisuje dva mechanismy epigenetické regulace v eukaryotách.
Klinické Zaměření: Travis, Usnesení
Tento příklad závěru Travis je příběh, který začal v Funkce Genetického Materiálu RNA, Transkripce, a Jak Asexuální Prokaryot Dosáhnout Genetické Rozmanitosti.,
přestože Travis přežil svůj zápas s nekrotizující fasciitidou, musel by nyní podstoupit operaci roubování kůže, následovanou dlouhodobou fyzikální terapií. Na základě množství svalové hmoty, kterou ztratil, je nepravděpodobné, že se jeho noha vrátí do plné síly, ale jeho fyzioterapeut je optimistický, že znovu využije nohu.
laboratorní vyšetření odhalilo, že původcem Travisovy infekce byl kmen streptokoka skupiny a (skupina a strep)., Jak vyžaduje zákon, Travisův případ byl hlášen ministerstvu zdravotnictví a nakonec centrům pro kontrolu a prevenci nemocí (CDC). Na CDC byl kmen skupiny a strep izolovaný od Travise důkladněji analyzován na rezistenci na meticilin.
rezistence na meticilin je geneticky zakódována a je stále častější ve skupině a strep prostřednictvím horizontálního přenosu genů. Při nekrotizující fasciitidě je průtok krve do infikované oblasti obvykle omezen kvůli působení různých geneticky kódovaných bakteriálních toxinů., To je důvod, proč je obvykle málo až žádné krvácení v důsledku řezu testu. Bohužel tyto bakteriální toxiny omezují účinnost intravenózních antibiotik při odstraňování infekce z kůže a podkladové tkáně, což znamená, že samotná rezistence na antibiotika nevysvětluje neúčinnost Travisovy léčby. Přesto byla zaručena intravenózní antibiotická terapie, která pomohla minimalizovat možný výsledek sepse, což je běžný výsledek nekrotizující fasciitidy., Prostřednictvím genomické analýzy CDC kmene izolovány od Travis, několik z důležitých virulentních genů bylo prokázáno, že být kódovány na prophages, což naznačuje, že transdukce je důležité v horizontální genový přenos těchto genů z jedné bakteriální buňky do druhé.
klíčové pojmy a souhrn
- genová exprese je přísně regulovaný proces.
- genová exprese v prokaryotách je do značné míry regulována v místě transkripce. Genová exprese v eukaryotách je dodatečně regulována po transkripci.,
- prokaryotické strukturní geny související funkce jsou často organizovány do operonů, vše je řízeno transkripcí z jediného promotoru. Regulační oblast operonu zahrnuje samotný promotor a oblast obklopující promotor, na kterou se transkripční faktory mohou vázat na ovlivnění transkripce.
- ačkoli jsou některé operony konstitutivně vyjádřeny, většina z nich podléhá regulaci pomocí transkripčních faktorů (represorů a aktivátorů)., A repressor se váže na operátor, sekvenci DNA v regulační oblasti mezi RNA polymeráza vazebné místo v promotoru a první strukturální gen, tím fyzicky blokují transkripci těchto operons. Aktivátor se váže v regulační oblasti operonu a pomáhá RNA polymeráze vázat se na promotor, čímž zvyšuje transkripci tohoto operonu. Induktor ovlivňuje transkripci interakcí s represorem nebo aktivátorem.
- operón trp je klasickým příkladem represivního operonu., Když se tryptofan hromadí, tryptofan se váže na represor, který se pak váže na operátora a zabraňuje další transkripci.
- lac operon je klasický příklad indukovatelný operon. Pokud je v buňce přítomna laktóza, převede se na alolaktózu. Allolaktóza působí jako induktor, který se váže na represor a zabraňuje vázání represoru na obsluhu. To umožňuje transkripci strukturních genů.
- Lac operon je také aktivován., Když jsou hladiny glukózy vyčerpány, některé buněčné ATP se přemění na cAMP, který se váže na protein aktivátoru katabolitu (CAP). Komplex cAMP-CAP aktivuje transkripci Lac operonu. Když jsou hladiny glukózy vysoké, jeho přítomnost zabraňuje transkripci Lac operonu a dalších opronů katabolitovou represí.
- malé intracelulární molekuly nazývané alarmony jsou vyráběny v reakci na různé environmentální stresy, což umožňuje bakteriím kontrolovat transkripci skupiny operanů, nazývaných regulon.,
- Bakterie mají schopnost měnit která σ faktor RNA polymerázy používají v reakci na podmínky životního prostředí, jak rychle a globálně změnit, které regulons jsou přepisována.
- Prokaryot mají regulační mechanismy, včetně útlumu a použití riboswitches, současně ovládat dokončení přepis a překlad z tohoto přepisu. Tyto mechanismy pracují prostřednictvím tvorby kmenových smyček na 5 ‚ konci molekuly mRNA, která je v současné době syntetizována.,
- existují další body regulace genové exprese u prokaryot a eukaryot. V eukaryotách je epigenetická regulace chemickou modifikací DNA nebo histonů a regulace zpracování RNA dvě metody.
Multiple Choice
operon genů kódujících enzymy v biosyntetické dráze je pravděpodobně který z následujících?
- indukovatelných
- repressible
- základní
- monocistronic
operon kódující geny, které jsou transkribovány a přeloženy nepřetržitě, aby poskytly buňce konstantní mezilehlé hladiny proteinových produktů, se říká, že jsou které z následujících?
- repressible
- indukovatelných
- základní
- aktivováno
která z následujících podmínek vede k maximálnímu vyjádření Lac operonu?
- laktóza přítomna, glukóza chybí
- laktóza přítomna, glukózy,
- laktóza chybí, glukózy chybí
- laktóza chybí, glukózy,
který z následujících je typ regulace genové exprese jedinečný pro eukaryoty?,
- útlum
- použít alternativní σ faktor
- chemické modifikace histonů
- alarmones
Vyplnit Prázdné
sekvence DNA, které repressors se může zavázat, že leží mezi promotorem a první strukturální gen se nazývá ________.,
prevence exprese operonů kódujících cesty substrátu pro substráty jiné než glukóza, pokud je přítomna glukóza, se nazývá_______.
- Jaké jsou dva způsoby, které bakterie mohou ovlivnit transkripci více různých operons současně v reakci na konkrétní stav životního prostředí?
- následující obrázek je z Monodovy původní práce na diauxickém růstu, který ukazuje růst E.coli v současné přítomnosti xylózy a glukózy jako jediného zdroje uhlíku. Vysvětlit, co se děje v bodech A–D s ohledem na zdroj uhlíku používány pro růst, a vysvětlit, zda xylózy-použití operon je vyjádřen (a proč)., Všimněte si, že exprese enzymů potřebných pro použití xylózy je regulována podobným způsobem jako exprese enzymů potřebných pro použití laktózy.
