Obiettivi formativi
- Confrontare operoni inducibili e operoni repressibili
- Descrivere perché la regolazione degli operoni è importante
Ogni cellula nucleata di un organismo multicellulare contiene copie dello stesso DNA. Allo stesso modo, tutte le cellule di due colture batteriche pure inoculate dalla stessa colonia iniziale contengono lo stesso DNA, ad eccezione dei cambiamenti che derivano da mutazioni spontanee., Se ogni cellula in un organismo multicellulare ha lo stesso DNA, allora come è possibile che le cellule in diverse parti del corpo dell’organismo mostrino caratteristiche diverse? Allo stesso modo, come è possibile che le stesse cellule batteriche all’interno di due colture pure esposte a diverse condizioni ambientali possano presentare fenotipi diversi? In entrambi i casi, ogni cellula geneticamente identica non accende o esprime lo stesso insieme di geni. Viene espresso solo un sottoinsieme di proteine in una cellula in un dato momento.,
Il DNA genomico contiene sia geni strutturali, che codificano prodotti che fungono da strutture cellulari o enzimi, sia geni regolatori, che codificano prodotti che regolano l’espressione genica. L’espressione di un gene è un processo altamente regolato. Mentre la regolazione dell’espressione genica negli organismi multicellulari consente la differenziazione cellulare, negli organismi unicellulari come i procarioti, garantisce principalmente che le risorse di una cellula non vengano sprecate producendo proteine di cui la cellula non ha bisogno in quel momento.,
Chiarire i meccanismi che controllano l’espressione genica è importante per la comprensione della salute umana. Malfunzionamenti in questo processo negli esseri umani portano allo sviluppo di cancro e altre malattie. Comprendere l’interazione tra l’espressione genica di un agente patogeno e quella del suo ospite umano è importante per la comprensione di una particolare malattia infettiva. La regolazione genica comporta una complessa rete di interazioni all’interno di una data cellula tra i segnali provenienti dall’ambiente cellulare, le molecole di segnalazione all’interno della cellula e il DNA della cellula., Queste interazioni portano all’espressione di alcuni geni e alla soppressione di altri, a seconda delle circostanze.
Procarioti ed eucarioti condividono alcune somiglianze nei loro meccanismi per regolare l’espressione genica; tuttavia, l’espressione genica negli eucarioti è più complicata a causa della separazione temporale e spaziale tra i processi di trascrizione e traduzione., Pertanto, sebbene la maggior parte della regolazione dell’espressione genica avvenga attraverso il controllo trascrizionale nei procarioti, la regolazione dell’espressione genica negli eucarioti avviene a livello trascrizionale e post-trascrizionale (dopo che è stata effettuata la trascrizione primaria).
Regolazione genica procariotica
Nei batteri e archaea, le proteine strutturali con funzioni correlate sono solitamente codificate insieme all’interno del genoma in un blocco chiamato operone e sono trascritte insieme sotto il controllo di un singolo promotore, con conseguente formazione di una trascrizione policistronica (Figura 1)., In questo modo, la regolazione della trascrizione di tutti i geni strutturali che codificano gli enzimi che catalizzano i molti passaggi in un singolo percorso biochimico può essere controllata simultaneamente, perché saranno tutti necessari allo stesso tempo, o nessuno sarà necessario. Ad esempio, in E. coli, tutti i geni strutturali che codificano gli enzimi necessari per utilizzare il lattosio come fonte di energia si trovano uno accanto all’altro nell’operone lattosio (o lac) sotto il controllo di un singolo promotore, il promotore lac., Gli scienziati francesi François Jacob (1920-2013) e Jacques Monod dell’Istituto Pasteur sono stati i primi a mostrare l’organizzazione dei geni batterici in operoni, attraverso i loro studi sull’operone lac di E. coli. Per questo lavoro, hanno vinto il premio Nobel per la Fisiologia o la Medicina nel 1965. Sebbene i geni eucariotici non siano organizzati in operoni, gli operoni procariotici sono modelli eccellenti per conoscere la regolazione genica in generale. Ci sono alcuni gruppi di geni negli eucarioti che funzionano in modo simile agli operoni., Molti dei principi possono essere applicati ai sistemi eucariotici e contribuire alla nostra comprensione dei cambiamenti nell’espressione genica negli eucarioti che possono causare cambiamenti patologici come il cancro.

Figura 1. Nei procarioti, i geni strutturali della funzione correlata sono spesso organizzati insieme sul genoma e trascritti insieme sotto il controllo di un singolo promotore. La regione regolatoria dell’operone comprende sia il promotore che l’operatore. Se un repressore si lega all’operatore, i geni strutturali non verranno trascritti., In alternativa, gli attivatori possono legarsi alla regione normativa, migliorando la trascrizione.
Ogni operone include sequenze di DNA che influenzano la propria trascrizione; queste si trovano in una regione chiamata regione regolatoria. La regione regolatrice comprende il promotore e la regione che circonda il promotore, a cui i fattori di trascrizione, proteine codificate da geni regolatori, possono legarsi. I fattori di trascrizione influenzano il legame della RNA polimerasi al promotore e consentono alla sua progressione di trascrivere i geni strutturali., Un repressore è un fattore di trascrizione che sopprime la trascrizione di un gene in risposta a uno stimolo esterno legandosi a una sequenza di DNA all’interno della regione regolatrice chiamata operatore, che si trova tra il sito di legame della RNA polimerasi del promotore e il sito di inizio trascrizionale del primo gene strutturale. Il legame del repressore blocca fisicamente la RNA polimerasi dalla trascrizione dei geni strutturali., Al contrario, un attivatore è un fattore di trascrizione che aumenta la trascrizione di un gene in risposta a uno stimolo esterno facilitando il legame della RNA polimerasi al promotore. Un induttore, un terzo tipo di molecola regolatrice, è una piccola molecola che attiva o reprime la trascrizione interagendo con un repressore o un attivatore.
Nei procarioti, ci sono esempi di operoni i cui prodotti genetici sono richiesti in modo piuttosto coerente e la cui espressione, quindi, non è regolamentata., Tali operoni sono espressi in modo costitutivo, nel senso che sono trascritti e tradotti continuamente per fornire alla cellula livelli intermedi costanti dei prodotti proteici. Tali geni codificano gli enzimi coinvolti nelle funzioni di pulizia necessarie per la manutenzione cellulare, compresa la replicazione del DNA, la riparazione e l’espressione, nonché gli enzimi coinvolti nel metabolismo del nucleo. Al contrario, ci sono altri operoni procarioti che sono espressi solo quando necessario e sono regolati da repressori, attivatori e induttori.,
Pensaci
- Quali sono le parti nella sequenza del DNA di un operone?
- Quali tipi di molecole regolatrici esistono?
Regolazione mediante repressione
Gli operoni procariotici sono comunemente controllati dal legame dei repressori alle regioni dell’operatore, impedendo così la trascrizione dei geni strutturali. Tali operoni sono classificati come operoni repressibili o operoni inducibili. Gli operoni repressibili, come l’operone del triptofano (trp), contengono tipicamente geni che codificano gli enzimi richiesti per una via biosintetica., Finché il prodotto della via, come il triptofano, continua ad essere richiesto dalla cellula, un operone reprimibile continuerà ad essere espresso. Tuttavia, quando il prodotto della via biosintetica inizia ad accumularsi nella cellula, rimuovendo la necessità che la cellula continui a produrre di più, l’espressione dell’operone viene repressa. Al contrario, gli operoni inducibili, come l’operone lac di E. coli, contengono spesso geni che codificano enzimi in una via coinvolta nel metabolismo di un substrato specifico come il lattosio., Questi enzimi sono richiesti soltanto quando quel substrato è disponibile, così l’espressione degli operoni è indotta tipicamente soltanto in presenza del substrato.
L’operone trp: Un operone reprimibile
E. coli può sintetizzare il triptofano usando enzimi codificati da cinque geni strutturali situati uno accanto all’altro nell’operone trp (Figura 2). Quando il triptofano ambientale è basso, l’operone è acceso. Ciò significa che la trascrizione è iniziata, i geni sono espressi e il triptofano è sintetizzato., Tuttavia, se il triptofano è presente nell’ambiente, l’operone trp è spento. La trascrizione non si verifica e il triptofano non viene sintetizzato.
Quando il triptofano non è presente nella cellula, il repressore da solo non si lega all’operatore; pertanto, l’operone è attivo e il triptofano viene sintetizzato. Tuttavia, quando il triptofano si accumula nella cellula, due molecole di triptofano si legano alla molecola del repressore trp, che cambia la sua forma, permettendole di legarsi all’operatore trp., Questo legame della forma attiva del repressore trp all’operatore blocca l’RNA polimerasi dalla trascrizione dei geni strutturali, fermando l’espressione dell’operone. Pertanto, il prodotto reale della via biosintetica controllata dall’operone regola l’espressione dell’operone.

Figura 2. I cinque geni strutturali necessari per sintetizzare il triptofano in E. coli si trovano uno accanto all’altro nell’operone trp. Quando il triptofano è assente, la proteina repressore non si lega all’operatore e i geni vengono trascritti., Quando il triptofano è abbondante, il triptofano lega la proteina del repressore alla sequenza dell’operatore. Questo blocca fisicamente la RNA polimerasi dalla trascrizione dei geni di biosintesi del triptofano.
Guarda questo video per saperne di più sull’operone trp.
L’Operone lac: Un Operone inducibile
L’operone lac è un esempio di operone inducibile che è anche soggetto ad attivazione in assenza di glucosio (Figura 3)., L’operone lac codifica tre geni strutturali necessari per acquisire ed elaborare il lattosio disaccaride dall’ambiente, scomponendolo negli zuccheri semplici glucosio e galattosio. Affinché l’operone lac sia espresso, il lattosio deve essere presente. Questo ha senso per la cellula perché sarebbe energeticamente dispendioso creare gli enzimi per elaborare il lattosio se il lattosio non fosse disponibile.
In assenza di lattosio, il repressore lac è legato alla regione operatore dell’operone lac, impedendo fisicamente alla RNA polimerasi di trascrivere i geni strutturali., Tuttavia, quando il lattosio è presente, il lattosio all’interno della cellula viene convertito in allolattosio. L’allolattosio funge da molecola induttore, legandosi al repressore e cambiando la sua forma in modo che non sia più in grado di legarsi al DNA dell’operatore. La rimozione del repressore in presenza di lattosio consente alla RNA polimerasi di muoversi attraverso la regione dell’operatore e iniziare la trascrizione dei geni strutturali lac.
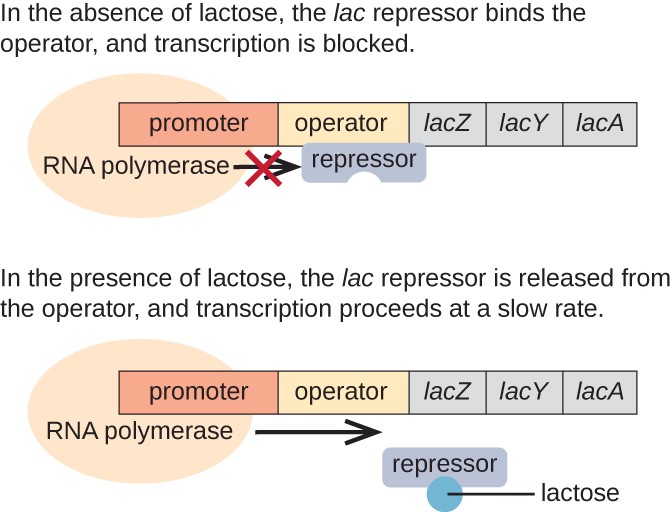
Figura 3. I tre geni strutturali necessari per degradare il lattosio in E. coli si trovano uno accanto all’altro nell’operone lac., Quando il lattosio è assente, la proteina repressore si lega all’operatore, bloccando fisicamente la RNA polimerasi dalla trascrizione dei geni strutturali lac. Quando il lattosio è disponibile, una molecola di lattosio lega la proteina repressore, impedendo al repressore di legarsi alla sequenza dell’operatore e i geni vengono trascritti.
L’operone lac: Attivazione da Catabolite Activator Protein

Figura 4. Se coltivato in presenza di due substrati, E. coli utilizza il substrato preferito (in questo caso glucosio) fino a quando non si esaurisce., Quindi, gli enzimi necessari per il metabolismo del secondo substrato vengono espressi e la crescita riprende, anche se a un ritmo più lento.
I batteri hanno tipicamente la capacità di utilizzare una varietà di substrati come fonti di carbonio. Tuttavia, poiché il glucosio è solitamente preferibile ad altri substrati, i batteri hanno meccanismi per garantire che i substrati alternativi vengano utilizzati solo quando il glucosio è stato esaurito. Inoltre, i batteri hanno meccanismi per garantire che i geni che codificano gli enzimi per l’utilizzo di substrati alternativi siano espressi solo quando il substrato alternativo è disponibile., Nel 1940, Jacques Monod è stato il primo a dimostrare la preferenza per alcuni substrati rispetto ad altri attraverso i suoi studi sulla crescita di E. coli quando coltivate in presenza di due substrati diversi contemporaneamente. Tali studi hanno generato curve di crescita diauxiche, come quella mostrata nella Figura 4. Sebbene il glucosio substrato preferito viene utilizzato per primo, E. coli cresce rapidamente e gli enzimi per il metabolismo del lattosio sono assenti. Tuttavia, una volta esauriti i livelli di glucosio, i tassi di crescita rallentano, inducendo l’espressione degli enzimi necessari per il metabolismo del secondo substrato, il lattosio., Si noti come il tasso di crescita del lattosio sia più lento, come indicato dalla minore pendenza della curva di crescita.
La capacità di passare dall’uso del glucosio ad un altro substrato come il lattosio è una conseguenza dell’attività di un enzima chiamato Enzima IIA (EIIA). Quando i livelli di glucosio diminuiscono, le cellule producono meno ATP dal catabolismo (vedi Catabolismo dei carboidrati) e l’EIIA diventa fosforilata., L’EIIA fosforilata attiva l’adenilil ciclasi, un enzima che converte parte dell’ATP rimanente in AMP ciclico (cAMP), un derivato ciclico dell’AMP e un’importante molecola di segnalazione coinvolta nel glucosio e nel metabolismo energetico in E. coli. Di conseguenza, i livelli del campo iniziano a salire nella cella (Figura 5).

Figura 5. Quando i livelli di ATP diminuiscono a causa dell’esaurimento del glucosio, alcuni ATP rimanenti vengono convertiti in cAMP dall’adenilil ciclasi. Pertanto, l’aumento dei livelli di cAMP segnala l’esaurimento del glucosio.,
L’operone lac svolge anche un ruolo in questo passaggio dall’uso del glucosio all’uso del lattosio. Quando il glucosio è scarso, l’accumulo di CAMP causato da un aumento dell’attività dell’adenilil ciclasi si lega alla proteina attivatore catabolita (CAP), nota anche come proteina recettore cAMP (CRP). Il complesso si lega alla regione promotrice dell’operone lac (Figura 6). Nelle regioni regolatorie di questi operoni, un sito di legame CAP si trova a monte del sito di legame della RNA polimerasi nel promotore., Il legame del complesso CAP-cAMP a questo sito aumenta la capacità di legame della RNA polimerasi alla regione del promotore di avviare la trascrizione dei geni strutturali. Pertanto, nel caso dell’operone lac, affinché si verifichi la trascrizione, il lattosio deve essere presente (rimuovendo la proteina repressore lac) e i livelli di glucosio devono essere esauriti (consentendo il legame di una proteina attivante). Quando i livelli di glucosio sono alti, c’è una repressione catabolita degli operoni che codificano gli enzimi per il metabolismo di substrati alternativi., A causa dei bassi livelli di campo in queste condizioni, c’è una quantità insufficiente del complesso CAP-cAMP per attivare la trascrizione di questi operoni. Si veda la tabella 1 per una sintesi del regolamento dell’operone lac.

Figura 6. (a) In presenza di CAMP, CAP si lega ai promotori degli operoni, come l’operone lac, che codificano i geni per gli enzimi per l’uso di substrati alternativi. b) Affinché l’operone lac sia espresso, deve essere attivata mediante cAMP-CAP e rimosso il repressore lac dall’operatore.,
| Tabella 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
Pensaci
- Cosa influenza il legame del repressore dell’operone trp all’operatore?
- Come e quando è alterato il comportamento della proteina repressore lac?
- Oltre ad essere reprimibile, in quale altro modo è regolato l’operone lac?
Risposte globali dei procarioti
Nei procarioti, ci sono anche diversi livelli più alti di regolazione genica che hanno la capacità di controllare la trascrizione di molti operoni correlati contemporaneamente in risposta a un segnale ambientale., Un gruppo di operoni tutti controllati simultaneamente è chiamato regulon.
Alarmones
Quando percepiscono lo stress imminente, i procarioti alterano l’espressione di un’ampia varietà di operoni per rispondere in coordinazione. Lo fanno attraverso la produzione di alarmoni, che sono piccoli derivati nucleotidici intracellulari. Gli alarmoni cambiano quali geni sono espressi e stimolano l’espressione di specifici geni di risposta allo stress. L’uso di alarmoni per alterare l’espressione genica in risposta allo stress sembra essere importante nei batteri patogeni., Incontrando i meccanismi di difesa dell’ospite ed altre circostanze dure durante l’infezione, molti operoni che codificano i geni di virulenza sono upregulated in risposta alla segnalazione di alarmone. La conoscenza di queste risposte è fondamentale per essere in grado di comprendere appieno il processo di infezione di molti agenti patogeni e per lo sviluppo di terapie per contrastare questo processo.,
Fattori σ alternativi
Poiché la subunità σ della RNA polimerasi batterica conferisce specificità su quali promotori devono essere trascritti, alterare il fattore σ utilizzato è un altro modo per i batteri di cambiare rapidamente e globalmente quali reguloni vengono trascritti in un dato momento. Il fattore σ riconosce le sequenze all’interno di un promotore batterico, quindi diversi fattori σ riconosceranno ciascuno sequenze di promotori leggermente diverse., In questo modo, quando la cellula percepisce specifiche condizioni ambientali, può rispondere cambiando quale fattore σ esprime, degradando quello vecchio e producendo uno nuovo per trascrivere gli operoni che codificano i geni i cui prodotti saranno utili nella nuova condizione ambientale. Ad esempio, nei batteri sporulanti dei generi Bacillus e Clostridium (che includono molti agenti patogeni), un gruppo di fattori σ controlla l’espressione dei molti geni necessari per la sporulazione in risposta a segnali stimolanti la sporulazione.,
Pensaci
- Qual è il nome dato a una raccolta di operoni che può essere regolata come un gruppo?
- Che tipo di stimolo attiverebbe la trascrizione di un diverso fattore σ?
Ulteriori metodi di regolazione nei batteri: Attenuazione e riboswitch
Sebbene la maggior parte dell’espressione genica sia regolata a livello di inizio della trascrizione nei procarioti, esistono anche meccanismi per controllare contemporaneamente sia il completamento della trascrizione che la traduzione., Dalla loro scoperta, questi meccanismi hanno dimostrato di controllare il completamento della trascrizione e della traduzione di molti operoni procarioti. Poiché questi meccanismi collegano direttamente la regolazione della trascrizione e della traduzione, sono specifici dei procarioti, poiché questi processi sono fisicamente separati negli eucarioti.
Un tale sistema normativo è l’attenuazione, per cui le strutture secondarie del gambo-ciclo formate all’interno dell’estremità 5’ di un mRNA che è trascritto determinano se la trascrizione per completare la sintesi di questo mRNA accadrà e se questo mRNA sarà usato per traduzione., Oltre al meccanismo di repressione trascrizionale già discusso, l’attenuazione controlla anche l’espressione dell’operone trp in E. coli (Figura 7). La regione regolatoria dell’operone trp contiene una sequenza leader chiamata trpL tra l’operatore e il primo gene strutturale, che ha quattro tratti di RNA che possono basarsi l’uno con l’altro in diverse combinazioni. Quando un terminatore stem-loop forme, trascrizione termina, rilasciando RNA polimerasi dal mRNA., Tuttavia, quando un antiterminator stem-loop forme, questo impedisce la formazione del terminator stem-loop, così RNA polimerasi può trascrivere i geni strutturali.
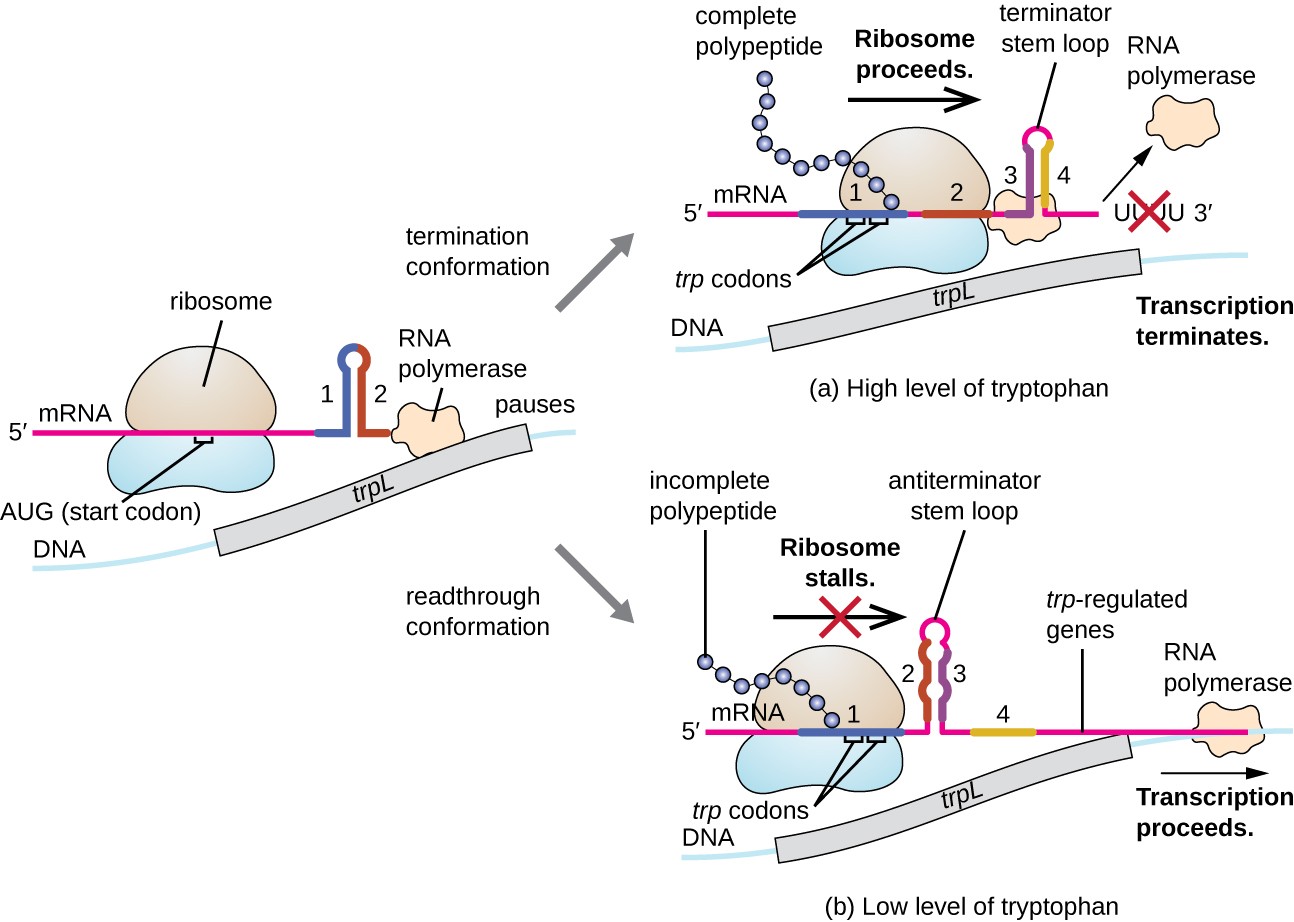
Figura 7. Fare clic per visualizzare un’immagine ingrandita. Quando il triptofano è abbondante, la traduzione del peptide breve del capo codificato da TRPL procede, il ciclo del terminatore fra le regioni 3 e 4 si forma e la trascrizione termina., Quando i livelli di triptofano sono esauriti, la traduzione del peptide leader corto si blocca nella regione 1, consentendo alle regioni 2 e 3 di formare un ciclo antiterminatore e la RNA polimerasi può trascrivere i geni strutturali dell’operone trp.
Un meccanismo correlato di regolazione simultanea della trascrizione e della traduzione nei procarioti è l’uso di un riboswitch, una piccola regione di RNA non codificante trovata all’interno dell’estremità 5’ di alcune molecole di mRNA procarioti (Figura 8). Un riboswitch può legarsi a una piccola molecola intracellulare per stabilizzare alcune strutture secondarie della molecola di mRNA., Il legame della piccola molecola determina quale struttura del gambo-ciclo si forma, influenzando così il completamento della sintesi dell’mRNA e della sintesi proteica.

Figura 8. Clicca per ingrandire l’immagine. I riboswitch trovati all’interno delle molecole di mRNA procarioti possono legarsi a piccole molecole intracellulari, stabilizzando alcune strutture di RNA, influenzando il completamento della sintesi della molecola di mRNA stessa (a sinistra) o la proteina prodotta usando quell’mRNA (a destra).,
Altri fattori che influenzano l’espressione genica nei procarioti e negli Eucarioti
Sebbene l’attenzione sulla nostra discussione sul controllo trascrizionale abbia usato come esempi gli operoni procarioti, il controllo trascrizionale eucariotico è simile in molti modi. Come nei procarioti, la trascrizione eucariotica può essere controllata attraverso il legame di fattori di trascrizione tra cui repressori e attivatori., È interessante notare che la trascrizione eucariotica può essere influenzata dal legame delle proteine alle regioni del DNA, chiamate enhancer, piuttosto lontane dal gene, attraverso il loop del DNA facilitato tra l’enhancer e il promotore (Figura 9). Nel complesso, la regolazione della trascrizione è un modo altamente efficace per controllare l’espressione genica sia nei procarioti che negli eucarioti. Tuttavia, il controllo dell’espressione genica negli eucarioti in risposta agli stress ambientali e cellulari può essere realizzato in modi aggiuntivi senza il legame dei fattori di trascrizione alle regioni regolatorie.,

Figura 9. Negli eucarioti, un potenziatore è una sequenza di DNA che promuove la trascrizione. Ogni potenziatore è costituito da brevi sequenze di DNA chiamate elementi di controllo distali. Gli attivatori legati agli elementi di controllo distali interagiscono con proteine mediatrici e fattori di trascrizione. Due geni diversi possono avere lo stesso promotore ma diversi elementi di controllo distale, consentendo l’espressione genica differenziale.,
Controllo a livello di DNA
Negli eucarioti, le molecole di DNA o gli istoni associati possono essere modificati chimicamente in modo da influenzare la trascrizione; questo è chiamato regolazione epigenetica. È stato dimostrato che la metilazione di alcuni nucleotidi della citosina nel DNA in risposta a fattori ambientali influenza l’uso di tale DNA per la trascrizione, con la metilazione del DNA comunemente correlata a livelli ridotti di espressione genica., Inoltre, in risposta a fattori ambientali, le proteine istoniche per il DNA di confezionamento possono anche essere modificate chimicamente in diversi modi, tra cui acetilazione e deacetilazione, influenzando lo stato di confezionamento del DNA e influenzando così la disponibilità di DNA vagamente avvolto per la trascrizione. Queste modifiche chimiche possono talvolta essere mantenute attraverso più cicli di divisione cellulare, rendendo ereditabili almeno alcuni di questi cambiamenti epigenetici.
Questo video descrive come la regolazione epigenetica controlla l’espressione genica.,
Pensaci
- Cosa interrompe o consente alla trascrizione di procedere quando l’attenuazione è in funzione?
- Cosa determina lo stato di un riboswitch?
- Descrivere la funzione di un potenziatore.
- Descrivere due meccanismi di regolazione epigenetica negli eucarioti.
Focus clinico: Travis, Resolution
Questo esempio conclude la storia di Travis iniziata nelle funzioni del materiale genetico, della trascrizione dell’RNA e di come i procarioti asessuati raggiungono la diversità genetica.,
Sebbene Travis sia sopravvissuto al suo attacco con fascite necrotizzante, ora dovrà sottoporsi a un intervento chirurgico di innesto cutaneo, seguito da una terapia fisica a lungo termine. Sulla base della quantità di massa muscolare che ha perso, è improbabile che la sua gamba ritorni a piena forza, ma il suo fisioterapista è ottimista sul fatto che recupererà un po ‘ di uso della sua gamba.
I test di laboratorio hanno rivelato che l’agente eziologico dell’infezione di Travis era un ceppo di streptococco di gruppo A (streptococco di gruppo A)., Come richiesto dalla legge, il caso di Travis è stato segnalato al dipartimento di salute dello stato e, infine, ai Centers for Disease Control and Prevention (CDC). Al CDC, il ceppo di streptococco di gruppo A isolato da Travis è stato analizzato più a fondo per la resistenza alla meticillina.
La resistenza alla meticillina è geneticamente codificata e sta diventando più comune nello streptococco di gruppo A attraverso il trasferimento genico orizzontale. Nella fascite necrotizzante, il flusso sanguigno verso l’area infetta è tipicamente limitato a causa dell’azione di varie tossine batteriche geneticamente codificate., Questo è il motivo per cui in genere c’è poco o nessun sanguinamento a seguito del test di incisione. Sfortunatamente, queste tossine batteriche limitano l’efficacia degli antibiotici per via endovenosa nell’eliminare l’infezione dalla pelle e dal tessuto sottostante, il che significa che la resistenza agli antibiotici da sola non spiega l’inefficacia del trattamento di Travis. Tuttavia, la terapia antibiotica endovenosa è stata giustificata per contribuire a minimizzare l’esito possibile della sepsi, che è un risultato comune della fascite necrotizzante., Attraverso l’analisi genomica da parte del CDC del ceppo isolato da Travis, molti degli importanti geni di virulenza hanno dimostrato di essere codificati su prophages, indicando che la trasduzione è importante nel trasferimento genico orizzontale di questi geni da una cellula batterica all’altra.
Concetti chiave e sintesi
- L’espressione genica è un processo strettamente regolato.
- L’espressione genica nei procarioti è in gran parte regolata al punto di trascrizione. L’espressione genica negli eucarioti è ulteriormente regolata post-trascrizionalmente.,
- I geni strutturali procariotici della funzione correlata sono spesso organizzati in operoni, tutti controllati dalla trascrizione da un singolo promotore. La regione regolatoria di un operone include il promotore stesso e la regione circostante il promotore a cui i fattori di trascrizione possono legarsi per influenzare la trascrizione.
- Sebbene alcuni operoni siano espressi in modo costitutivo, la maggior parte sono soggetti a regolazione attraverso l’uso di fattori di trascrizione (repressori e attivatori)., Un repressore si lega ad un operatore, una sequenza di DNA all’interno della regione regolatrice tra il sito di legame della RNA polimerasi nel promotore e il primo gene strutturale, bloccando così fisicamente la trascrizione di questi operoni. Un attivatore si lega all’interno della regione regolatrice di un operone, aiutando l’RNA polimerasi a legarsi al promotore, migliorando così la trascrizione di questo operone. Un induttore influenza la trascrizione interagendo con un repressore o attivatore.
- L’operone trp è un classico esempio di operone reprimibile., Quando il triptofano si accumula, il triptofano si lega a un repressore, che quindi si lega all’operatore, impedendo un’ulteriore trascrizione.
- L’operone lac è un classico esempio di un operone inducibile. Quando il lattosio è presente nella cellula, viene convertito in allolattosio. L’allolattosio agisce come un induttore, legandosi al repressore e impedendo al repressore di legarsi all’operatore. Ciò consente la trascrizione dei geni strutturali.
- Anche l’operone lac è soggetto ad attivazione., Quando i livelli di glucosio sono esauriti, alcuni ATP cellulare viene convertito in cAMP, che si lega alla proteina attivatore catabolita (CAP). Il complesso cAMP-CAP attiva la trascrizione dell’operone lac. Quando i livelli di glucosio sono alti, la sua presenza impedisce la trascrizione dell’operone lac e di altri operoni mediante repressione catabolita.
- Piccole molecole intracellulari chiamate alarmoni sono prodotte in risposta a vari stress ambientali, consentendo ai batteri di controllare la trascrizione di un gruppo di operoni, chiamato regulon.,
- I batteri hanno la capacità di cambiare quale σ fattore di RNA polimerasi usano in risposta alle condizioni ambientali per cambiare rapidamente e globalmente quali reguloni vengono trascritti.
- I procarioti hanno meccanismi regolatori, tra cui l’attenuazione e l’uso di riboswitch, per controllare simultaneamente il completamento della trascrizione e della traduzione da quella trascrizione. Questi meccanismi funzionano con la formazione dei cicli del gambo nell’estremità 5 ‘ di una molecola di mRNA attualmente che è sintetizzata.,
- Ci sono ulteriori punti di regolazione dell’espressione genica nei procarioti e negli eucarioti. Negli eucarioti, la regolazione epigenetica mediante modificazione chimica del DNA o degli istoni e la regolazione dell’elaborazione dell’RNA sono due metodi.
Scelta multipla
È probabile che un operone di geni che codificano enzimi in una via biosintetica sia quale dei seguenti?
- inducibile
- repressibile
- costitutivo
- monocistronico
Un operone codifica geni che vengono trascritti e tradotti continuamente per fornire la cellula con livelli intermedi costanti dei prodotti proteici è detto di essere quale dei seguenti?
- repressibile
- inducibile
- costitutivo
- attivato
Quale delle seguenti condizioni porta alla massima espressione dell’operone lac?
- il lattosio presente, glucosio assente
- il lattosio presente, il glucosio presente
- lattosio assenti glucosio assente
- lattosio assenti, il glucosio presente
Quale dei seguenti è un tipo di regolazione dell’espressione genica unica per gli eucarioti?,
- attenuazione
- uso alternativo σ fattore
- modifica chimica di istoni
- alarmones
Riempire il vuoto
La sequenza di DNA, a cui i repressori possono legarsi, che si trova tra il promotore e il primo gene strutturale è chiamata ________.,
La prevenzione dell’espressione degli operoni che codificano i percorsi di utilizzo del substrato per substrati diversi dal glucosio quando il glucosio è presente è chiamata _______.
Pensaci
- Quali sono due modi in cui i batteri possono influenzare la trascrizione di più operoni diversi contemporaneamente in risposta a una particolare condizione ambientale?
- La figura seguente è tratta dal lavoro originale di Monod sulla crescita diauxica che mostra la crescita di E. coli in presenza simultanea di xilosio e glucosio come uniche fonti di carbonio. Spiegare cosa sta succedendo nei punti A-D rispetto alla fonte di carbonio utilizzata per la crescita e spiegare se viene espresso l’operone per uso di xilosio (e perché)., Si noti che l’espressione degli enzimi necessari per l’uso di xilosio è regolata in modo simile all’espressione degli enzimi necessari per l’uso di lattosio.
