Mål
- Sammenlign inducible operons og repressible operons
- Beskrive hvorfor regulering av operons er viktig
Hver nucleated celle i en flercellede organismen inneholder kopier av samme DNA. På samme måte, alle celler i to rene bakteriekulturer inokulert fra samme starter koloni inneholder det samme DNA, med unntak av endringer som oppstår fra spontane mutasjoner., Hvis hver celle i en flercellede organismen har det samme DNA, så hvordan har det seg at celler i ulike deler av organismens kroppen har ulike egenskaper? På samme måte, hvordan er det at den samme bakteriell celler innen to rene kulturer utsatt for ulike miljøforhold kan ha forskjellige phenotypes? I begge tilfeller, hver genetisk identiske cellen ikke slå på, eller uttrykk, samme sett av gener. Bare et delsett av proteiner i en celle på et gitt tidspunkt er uttrykt.,
Genomisk DNA inneholder både strukturelle gener som koder for produkter som fungerer som cellulære strukturer eller enzymer, og regulatoriske gener som koder for produkter som regulerer genuttrykk. Uttrykket av et gen er en sterkt regulert prosess. Mens regulering av genuttrykk i flercellede organismer kan for cellulær differensiering, i encellede organismer som prokaryotes, er det først og fremst sørger for at en celle ‘ s ressurser er ikke bortkastet å lage proteiner som cellen trenger ikke på det tidspunktet.,
Elucidating de mekanismene som styrer genuttrykk er viktig for forståelsen av menneskers helse. Feil i denne prosessen hos mennesker fører til utvikling av kreft og andre sykdommer. Forstå samspillet mellom genuttrykk av en patogen og dets hærskarene som er viktig for forståelse av en bestemt smittsom sykdom. Genet reguleringen består av et komplisert nett av interaksjoner innenfor en gitt celle blant signaler fra cellens omgivelser, signal-molekyler i cellen, og cellens DNA., Disse vekselsvirkningene føre til uttrykk av enkelte gener og undertrykkelse av andre, avhengig av omstendighetene.
Prokaryotes og eukaryotes dele noen likheter i deres mekanismer for å regulere genuttrykk, men genuttrykk i eukaryotes er mer komplisert på grunn av den temporale og romlige avstanden mellom prosesser av transkripsjon og oversettelse., Dermed, selv om de fleste regulering av genuttrykk skjer gjennom transcriptional kontroll i prokaryotes, regulering av genuttrykk i eukaryotes skjer på transcriptional nivå og post-transcriptionally (etter den primære transkripsjonen er gjort).
Prokaryotic Gene Regulation
I bakterier og archaea, strukturelle proteiner med relaterte funksjoner er vanligvis kodet sammen i genomet i en blokk kalles en operon og er skrevet sammen under kontroll av en enkelt arrangøren, noe som resulterer i dannelsen av en polycistronic transkripsjonen (Figur 1)., På denne måten, regulering av transkripsjon av alle de strukturelle genene som koder enzymer som katalysere de mange trinn i en enkelt biokjemiske veien kan kontrolleres samtidig, fordi de vil alle være nødvendig på samme tid, eller ingen vil være nødvendig. For eksempel, i E. coli, alle strukturelle gener som koder for enzymer som trengs for å bruke laktose som energikilde ligge ved siden av hverandre i den laktose (eller lac) operon under kontroll av en enkelt arrangøren, den lac arrangøren., Franske forskere François Jakob (1920-2013) og Jacques Monod ved Pasteur-Instituttet ble den første til å vise organiseringen av bakterielle gener i operons, gjennom sine studier av lac operon av E. coli. For dette arbeidet, de vant nobelprisen i Fysiologi eller Medisin i 1965. Selv om eukaryote gener er ikke organisert i operons, prokaryotic operons er gode modeller for å lære om genet regulering generelt. Det er noen genet klynger i eukaryotes at funksjonen ligner operons., Mange av prinsippene kan brukes til eukaryote systemer og bidra til vår forståelse av endringer i genuttrykk i eukaryotes som kan føre til patologiske forandringer, for eksempel kreft.

Figur 1. I prokaryotes, strukturelle gener av beslektet funksjon er ofte organisert sammen på genom-og transkribert sammen under kontroll av en enkelt søker. Den operon er regulatoriske området omfatter både arrangøren og operatør. Hvis en repressor binder seg til operatøren, så det strukturelle gener vil ikke bli transkribert., Alternativt, utløsere kan binde seg til regulatoriske regionen, styrke transkripsjon.
Hver operon inkluderer DNA-sekvenser som har innflytelse på sin egen transkripsjon; disse ligger i et område kalt det regulatoriske området. Det regulatoriske området omfatter selskapet og regionen rundt selskapet, som transkripsjonfaktorer, proteiner kodet av regulatoriske gener, kan binde seg. Transkripsjon faktorer påvirker binding av RNA polymerase arrangøren og la dens progresjon å transkribere strukturelle gener., En repressor er en transkripsjonsfaktor som undertrykker transkripsjon av et gen i respons til en ekstern stimulans ved binding til DNA-sekvensen i det regulatoriske området kalt operatøren, som ligger mellom RNA polymerase bindende nettstedet av arrangøren og transcriptional start siden av den første strukturelle genet. Repressor bindende fysisk blokkerer RNA-polymerase fra transkribere strukturelle gener., I motsatt fall, en aktivator er en transkripsjonsfaktor som øker transkripsjon av et gen i respons til en ekstern stimulans ved å legge til rette RNA polymerase binding til arrangøren. En induser, en tredje type av regulatoriske molekyl, som er et lite molekyl som enten aktiverer eller undertrykker transkripsjon ved å samhandle med en repressor eller en aktivator.
I prokaryotes, det er eksempler på operons som gen-produkter må heller konsekvent og som uttrykk, derfor er uregulert., Slik operons er constitutively uttrykt, noe som betyr at de er transkribert og oversatt kontinuerlig for å gi cellen med konstant mellomliggende nivåer av protein-produkter. Slike gener som koder for enzymer som er involvert i rengjøring funksjonene som kreves for mobil vedlikehold, inkludert DNA-replikasjon, reparasjon og uttrykk, samt enzymer som er involvert i kjernen metabolismen. I motsetning, det er andre prokaryotic operons som er uttrykt bare når det er nødvendig og er regulert av repressors, utløsere, og indusere.,
Tenk på Det
- Hva er de delene i DNA sekvens av en operon?
- Hvilke typer av regulatoriske molekyler er det?
Reguleringen av Undertrykkelse
Prokaryotic operons er ofte styrt av binding av repressors til operatøren regioner, og dermed hindre transkripsjon av den strukturelle gener. Slik operons er klassifisert som enten repressible operons eller inducible operons. Repressible operons, som tryptofan (trp) operon, vanligvis inneholder genene som koder for enzymer som er nødvendige for en biosynthetic veien., Så lenge produktet av veien, som tryptofan, fortsetter å være nødvendig av cellen, en repressible operon vil fortsette å bli uttrykt. Imidlertid, når produktet av biosynthetic veien begynner å hope seg opp i cellen, noe som eliminerer behovet for cellen til å fortsette å gjøre mer, uttrykk for operon er undertrykt. I motsatt fall, inducible operons, som lac operon av E. coli, inneholder ofte genene som koder for enzymer i en sti som er involvert i metabolismen av en bestemt substrat som laktose., Disse enzymene er bare nødvendig når underlaget er tilgjengelige, og dermed uttrykk for operons er vanligvis forårsaket bare i nærvær av underlaget.
De trp Operon: En Repressible Operon
E. coli kan syntetisere tryptofan ved hjelp av enzymer som er kodet av fem strukturelle gener ligger ved siden av hverandre i trp operon (Figur 2). Når miljø-tryptofan er lav, operon er slått på. Dette betyr at transkripsjon er i gang, gener er uttrykt, og tryptofan er syntetisert., Imidlertid, hvis tryptofan er til stede i miljøet, trp operon er slått av. Transkripsjon ikke finne sted, og tryptofan er ikke syntetisk.
Når tryptofan er ikke til stede i cellen, repressor av seg selv binder seg ikke til operatøren; derfor, operon er aktiv og tryptofan er syntetisert. Imidlertid, når tryptofan akkumuleres i cellen, to tryptofan molekylene binder seg til trp repressor molekyl, som skifter form, slik at det å binde seg til trp operatør., Denne binding av den aktive formen av trp repressor til operatøren blokker RNA-polymerase fra transkribere de strukturelle gener, stopper uttrykk for operon. Dermed er den faktiske produktet av biosynthetic vei kontrollert av operon regulerer uttrykk av operon.

Figur 2. De fem strukturelle gener som trengs for å syntetisere tryptofan i E. coli er plassert ved siden av hverandre i trp operon. Når tryptofan er borte, repressor protein binder seg ikke til operatør, og genene er transkribert., Når tryptofan er rikelig, tryptofan binder repressor protein på operatør-sekvensen. Dette er fysisk blokkerer RNA-polymerase fra transkribere den tryptofan biosyntese gener.
Se denne videoen for å lære mer om trp operon.
De lac Operon: En Inducible Operon
lac operon er et eksempel på en inducible operon som også er gjenstand for aktivering i fravær av glukose (Figur 3)., Den lac operon koder tre strukturelle gener som er nødvendig for å tilegne seg og behandle disaccharide laktose fra miljøet, å bryte det ned til enkle sukkerarter som glukose og galaktose. For lac operon til å bli uttrykt, laktose må være til stede. Dette er fornuftig for cellen fordi det ville være energisk unødvendig å lage enzymer til å behandle laktose hvis laktose ikke var tilgjengelig.
I fravær av laktose, det lac repressor er bundet til operatøren region av lac operon, fysisk å hindre RNA-polymerase fra transkribere de strukturelle gener., Imidlertid, når laktose er til stede, laktose inne i cellen er konvertert til allolactose. Allolactose fungerer som en induser molekyl, binding til repressor og endre sin form, slik at det ikke lenger er i stand til å binde seg til operatøren DNA. Fjerning av repressor i nærvær av laktose kan RNA polymerase til å flytte gjennom operatøren regionen og begynner i transkripsjon av lac strukturelle gener.
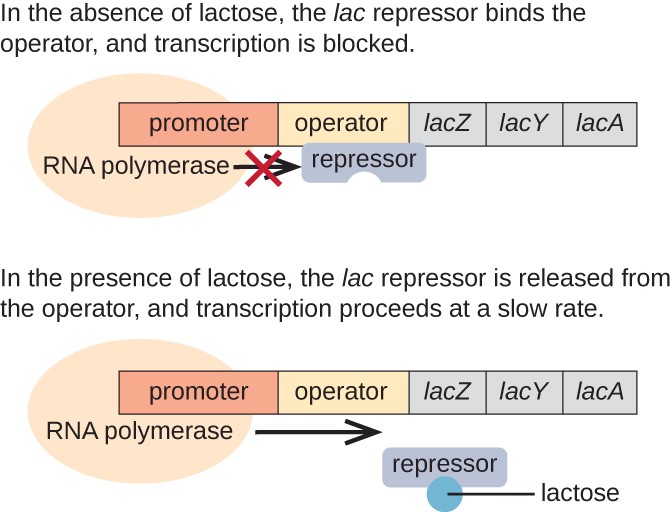
Figur 3. De tre strukturelle gener som er nødvendig for å bryte ned laktose i E. coli er plassert ved siden av hverandre i lac-operon., Når laktose er borte, repressor protein som binder seg til operatør, fysisk blokkere RNA-polymerase fra transkribere den lac strukturelle gener. Når laktose er tilgjengelig, laktose molekyl som binder repressor protein, hindrer repressor fra binding til operatøren rekkefølge, og genene er transkribert.
De lac Operon: Aktivering av Catabolite Aktivator Protein

Figur 4. Når vokst i nærvær av to underlag, E. coli bruker foretrukket substrat (i dette tilfellet glukose) til det er tomt., Så, enzymer som er nødvendig for metabolismen av andre underlaget er uttrykt, og veksten fortsetter, men med en lavere pris.
vanligvis Bakterier har evnen til å bruke et utvalg av underlag som karbon kilder. Imidlertid, fordi glukose er vanligvis å foretrekke fremfor andre substrater, bakterier har mekanismer for å sikre at alternativ underlag er kun brukt når glukose har blitt utarmet. I tillegg, bakterier har mekanismer for å sikre at genene som koder for enzymer for å bruke alternative underlag er uttrykt bare når den alternative underlaget er tilgjengelig., I 1940-årene, Jacques Monod var den første til å demonstrere preferanse for visse underlag over andre gjennom sine studier av E. coli vekst når kultivert i nærvær av to forskjellige underlag samtidig. Slike studier generert diauxic vekstkurver, som vist i Figur 4. Selv om foretrukket substrat glukose er brukt først, E. coli vokser raskt og enzymer for laktose metabolisme er fraværende. Men, når blodsukkeret er utladet, vekst sakte, inducing uttrykk for enzymer som er nødvendig for metabolismen av andre substrat, laktose., Legg merke til hvordan veksten i laktose er tregere, som indikert av lavere steepness av vekstkurve.
evnen til å bytte fra glukose bruk til et annet substrat som laktose er en konsekvens av aktiviteten til et enzym som kalles Enzym IIA (EIIA). Når glukose nivåer slipp, celler produserer mindre ATP fra katabolisme (se Nedbrytningen av Karbohydrater), og EIIA blir phosphorylated., Phosphorylated EIIA aktiverer adenylyl cyclase, et enzym som omdanner noen av de gjenværende ATP til syklisk AMP (cAMP), en syklisk derivat av FORSTERKER og viktig signal molekyl som er involvert i glukose-og energiomsetningen i E. coli. Som et resultat, cAMP nivåer begynner å stige i cellen (Figur 5).

Figur 5. Når ATP nivåer redusere på grunn av nedbryting av glukose, og noen gjenværende ATP er konvertert til cAMP ved adenylyl cyclase. Dermed auka cAMP nivå signal glukose-forbruk.,
lac operon spiller også en rolle i dette bytte fra å bruke glukose til å bruke laktose. Når glukose er knappe, accumulating cAMP forårsaket av økt adenylyl cyclase aktivitet binder seg til catabolite aktivator protein (CAP), også kjent som cAMP reseptor protein (CRP). Komplekset binder seg til arrangøren region av lac operon (Figur 6). I den regulatoriske regioner av disse operons, en CAP bindende nettstedet ditt er plassert oppstrøms av RNA polymerase bindende nettstedet i selskapet., Binding av CAP-cAMP komplekse til dette nettstedet øker bindende evne av RNA polymerase arrangøren regionen for å starte transkripsjon av strukturelle gener. Derfor, i tilfelle av lac operon, for transkripsjon til å skje, laktose må være til stede (fjerning av lac repressor protein) og glukose nivåer må være tømt (slik at bindingen av en aktivering av protein). Når blodsukkeret er høyt, det er catabolite undertrykkelse av operons koding enzymer for metabolismen av alternative underlag., På grunn av lave cAMP nivåer under disse forholdene, det er en utilstrekkelig mengde av CAP-cAMP komplisert å aktivere transkripsjon av disse operons. Se Tabell 1 for en oversikt over regulering av lac operon.

Figur 6. (a) I nærvær av leiren, CAP binder seg til arrangører av operons, som lac operon, som kodes av gener for enzymer for bruk av alternative underlag. (b) For lac operon å komme til uttrykk, må det være aktivering av cAMP-CAP, samt fjerning av lac repressor fra operatøren.,
| Tabell 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
Tenk på Det
- Hva påvirker binding av trp operon repressor til operatør?
- Hvordan og når er oppførselen til lac repressor protein endret?
- I tillegg til å være repressible, hvordan ellers er lac operon regulert?
Globale Tiltak av Prokaryotes
I prokaryotes, det er også flere høyere nivåer av genet, regulering som har muligheten til å kontrollere transkripsjon av mange i slekt operons samtidig i respons til en miljø-signal., En gruppe av operons alle kontrollert samtidig er kalt en regulon.
Alarmones
Når sensing forestående stress, prokaryotes endre uttrykket av et bredt utvalg av operons til å svare på koordinering. De gjør dette gjennom produksjon av alarmones, som er små intracellulære nukleotid derivater. Alarmones endre hvilke gener som blir uttrykt, og stimulere til uttrykket av spesifikke stress-respons-gener. Bruk av alarmones å endre genuttrykk i respons til stress ser ut til å være viktig i patogene bakterier., På møtet vert forsvarsmekanismer og andre tøffe forhold under infeksjonen, mange operons koding virulence gener er oppregulert i respons til alarmone signalering. Kunnskap om disse svarene er nøkkelen til å være i stand til å fullt ut forstå infeksjon prosess av mange patogener og til utvikling av terapi for å motvirke denne prosessen.,
Alternativ σ Faktorer
Siden σ subunit av bakterielle RNA polymerase gir spesifisitet som arrangører skulle bli transkribert, endre σ faktor som brukes er en annen måte for bakterier å raskt og globalt endre hva regulons er transkribert på et gitt tidspunkt. Den σ faktor gjenkjenner sekvenser i en bakteriell arrangøren, så forskjellige σ faktorer vil hver gjenkjenne litt annerledes arrangøren sekvenser., På denne måten, når cellen sanser spesifikke miljømessige forhold, kan den reagere ved å endre hvor σ faktor det uttrykker, nedverdigende den gamle og produsere en ny en for å transkribere den operons koding gener som produkter som vil være nyttige under den nye miljøbetingelser. For eksempel, i sporulating bakterier av slektene Bacillus og Clostridium (som inkluderer mange patogener), en gruppe av σ faktorer som styrer uttrykket av mange gener som trengs for sporulation i respons til sporulation-stimulerende signaler.,
Tenk på Det
- Hva er navnet gitt til en samling av operons som kan bli regulert i en gruppe?
- Hvilken type stimulus ville utløse transkripsjon av en annen σ faktor?
Flere Metoder for Regulering i Bakterier: Demping og Riboswitches
Selv om de fleste genuttrykk reguleres på nivå med transkripsjon innvielse i prokaryotes, det er også mekanismer for å styre både gjennomføring av transkripsjon, samt oversettelse samtidig., Siden oppdagelsen deres, disse mekanismene har vist seg å styre gjennomføring av transkripsjon og oversettelse av mange prokaryotic operons. Fordi disse mekanismene link regulering av transkripsjon og oversettelse direkte, de er spesifikke for prokaryotes, fordi disse prosessene er fysisk adskilt i eukaryotes.
Et slikt reguleringssystem er demping, der sekundære stilk-loop strukturer dannet i 5′ – enden av en mRNA blir transkribert avgjøre om transkripsjon for å fullføre syntese av dette mRNA vil oppstå, og hvis dette mRNA vil bli brukt for oversettelse., Utover transcriptional undertrykkelse mekanisme som allerede diskutert, demping styrer også uttrykk for trp operon i E. coli (Figur 7). Den trp operon regulatoriske området inneholder en leder sekvens som kalles trpL mellom operatør og den første strukturelle genet, som har fire strekninger av RNA som kan basere par med hverandre i forskjellige kombinasjoner. Når en terminator-stem-loop former, avslutter transkripsjonen, slippe RNA-polymerase fra mRNA., Men, når en antiterminator stilk-loop former, dette forhindrer dannelsen av terminator-stem-loop, så RNA polymerase kan transkribere de strukturelle gener.
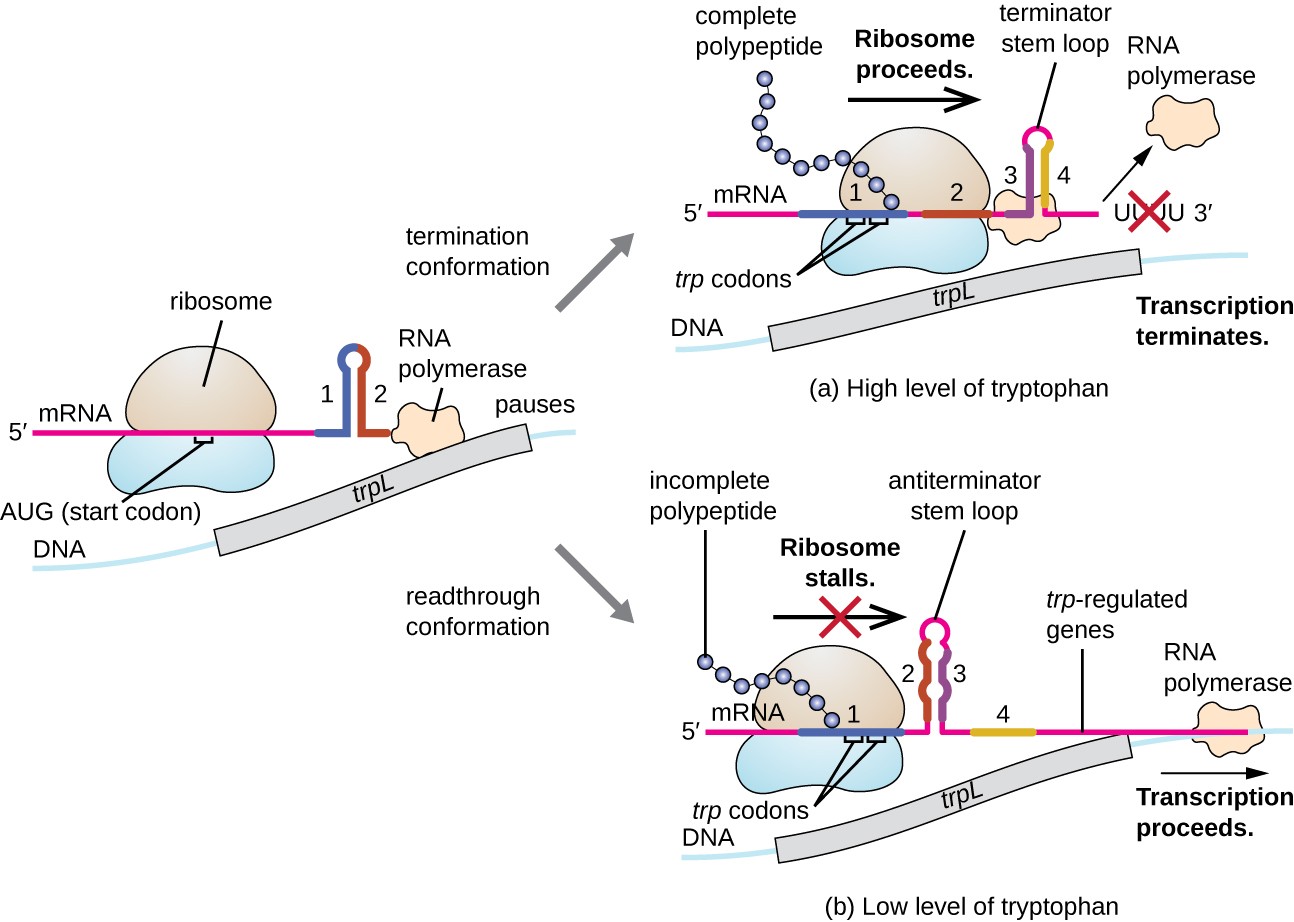
Figur 7. Klikk for å se et større bilde. Når tryptofan er rikelig, oversettelse av den korte leder peptid kodet av trpL inntektene, terminator loop mellom regioner 3 og 4 former, og transkripsjon opphører., Når tryptofan nivåer er oppbrukt, oversettelse av den korte leder peptid boder i region 1, slik at regionene 2 og 3 for å danne en antiterminator loop -, og RNA-polymerase kan transkribere de strukturelle gener av trp operon.
En relatert mekanisme samtidige regulering av transkripsjon og oversettelse i prokaryotes er bruken av en riboswitch, et lite område av noncoding RNA finnes i 5′ – enden av noen prokaryotic mRNA-molekyler (Figur 8). En riboswitch kan binde seg til en liten intracellulære molekyl til å stabilisere visse sekundære strukturer av mRNA-molekyl., Binding av små molekyl bestemmer hvilken stamme-løkke struktur former, og dermed påvirke gjennomføring av mRNA-syntese og proteinsyntese.

Figur 8. Klikk for større bilde. Riboswitches funnet i løpet av prokaryotic mRNA-molekyler som kan binde seg til små intracellulære molekyler, stabiliserende enkelte RNA-strukturer, som påvirker enten ferdigstillelse av syntese av mRNA-molekylet seg selv (venstre) eller protein som er gjort med at mRNA (høyre).,
Andre Faktorer som Påvirker genuttrykk i Prokaryotes og Eukaryotes
Selv om fokus på vår diskusjon av transcriptional kontroll brukt prokaryotic operons som eksempler, eukaryote transcriptional kontroll er lik på mange måter. Som i prokaryotes, eukaryote transkripsjon kan være kontrollert gjennom binding av transkripsjon faktorer, inkludert repressors og utløsere., Interessant, eukaryote transkripsjon kan være påvirket av binding av proteiner til områder av DNA, kalt enhancers, ganske langt bort fra genet, gjennom DNA-løkker tilrettelagt mellom enhancer og arrangøren (Figur 9). Totalt sett, regulering av transkripsjon er en svært effektiv måte å kontrollere genuttrykk i både prokaryotes og eukaryotes. Imidlertid, kontroll av genuttrykk i eukaryotes i respons til miljø-og cellulære stress kan oppnås på flere måter, uten binding av transkripsjonfaktorer til regulatoriske regioner.,

Figur 9. I eukaryotes, en enhancer er en DNA-sekvens som fremmer transkripsjon. Hver enhancer er bygd opp av korte DNA-sekvenser som kalles distale betjeningselementer. Aktivatorene bundet til den distale kontroll elementene samhandle med mekleren proteiner og transkripsjon faktorer. To ulike gener kan ha samme arrangøren, men forskjellige distale betjeningselementer, som muliggjør differensiert genuttrykk.,
DNA-Nivå Kontroll
I eukaryotes, DNA-molekyler eller forbundet histones kan være kjemisk modifisert på en slik måte som å påvirke transkripsjon; dette kalles epigenetisk regulering. Metylering av visse cytosine nukleotider i DNA i respons til miljømessige faktorer har vist seg å påvirke bruken av slike DNA for transkripsjon, med DNA-metylering vanlig å sammenstille til reduserte nivåer av genuttrykk., I tillegg, i respons på miljøfaktorer, histone proteiner for emballasje DNA kan også være kjemisk endret på flere måter, blant annet acetylation og deacetylation, påvirke emballasje tilstand av DNA, og dermed påvirke tilgjengeligheten av løst såret DNA for transkripsjon. Disse kjemiske modifikasjoner kan noen ganger være vedlikeholdt gjennom flere runder med celledeling, noe som gjør minst noen av disse epigenetisk endringer heritable.
Denne videoen beskriver hvordan epigenetisk regulering styrer genuttrykk.,
Tenk på Det
- Hva stopper eller gjør transkripsjon for å gå videre når demping fungerer?
- Hva bestemmer staten en riboswitch?
- Beskrive funksjonen til en enhancer.
- Beskrive to mekanismer for epigenetisk regulering i eukaryotes.
Klinisk Fokus: Travis, Oppløsning
Dette eksemplet konkluderer med Travis ‘ s historie som startet i Funksjonene av Genetisk Materiale, RNA-Transkripsjon, og Hvordan Asexual Prokaryotes Oppnå Genetisk Mangfold.,
Selv om Travis overlevde hans kamp med necrotizing fasciitis, ville han nå har til å gjennomgå en hud-pode kirurgi, etterfulgt av langvarig fysisk terapi. Basert på mengde muskelmasse han mistet, er det lite sannsynlig at benet vil komme tilbake til full styrke, men hans fysioterapeut er optimistisk i forhold til at han vil få noen bruk av benet.
Laboratorium tester viste causative agent for Travis ‘ s infeksjon ble en belastning av gruppe A-streptokokker (Gruppe A strep)., Det som kreves av loven, Travis er saken ble rapportert til statens helse avdeling og til slutt til Centers for Disease Control og Prevention (CDC). På CDC, belastningen av gruppe A strep isolert fra Travis ble analysert mer grundig for meticillin motstand.
Meticillin-motstand er genetisk kodet og blir mer og mer vanlig i gruppe A strep gjennom horisontal gene transfer. I necrotizing fasciitis, blodtilførselen til det infiserte området er vanligvis begrenset på grunn av virkningen av ulike genetisk kodet bakterielle toksiner., Dette er hvorfor det er vanligvis lite eller ingen blødninger som følge av snittet test. Dessverre, disse bakterielle toksiner begrense effektiviteten av intravenøs antibiotika i å fjerne infeksjonen fra hud og underliggende vev, noe som betyr at antibiotika resistens alene ikke kan forklare ineffektivitet av Travis ‘ s behandling. Likevel, intravenøs antibiotikabehandling var berettiget til å minimere mulige utfall av sepsis, som er et vanlig utfall av necrotizing fasciitis., Gjennom genomisk analyse av CDC av belastningen isolert fra Travis, flere av de viktige virulence gener ble vist å være kodet på prophages, noe som indikerer at transduksjon er viktig i den horisontale genet overføring av disse gener fra en bakterie celle til en annen.
sentrale Begreper og Oppsummering
- genuttrykk er en strengt regulert prosess.
- genuttrykk i prokaryotes er i stor grad regulert på det punktet av transkripsjon. Genuttrykk i eukaryotes er i tillegg regulert av post-transcriptionally.,
- Prokaryotic strukturelle gener av beslektet funksjon er ofte organisert i operons, alle kontrollert av transkripsjon fra en enkelt søker. Det regulatoriske området for en operon inkluderer arrangøren seg selv og regionen rundt arrangøren som transkripsjonfaktorer kan binde til å påvirke transkripsjon.
- Selv om noen operons er constitutively uttrykt, de fleste er underlagt regulering gjennom bruk av transkripsjon faktorer (repressors og utløsere)., En repressor binder seg til en operatør, en DNA-sekvens i det regulatoriske området mellom RNA polymerase bindende nettstedet i selskapet og første strukturelle gen, og dermed fysisk blokkere transkripsjon av disse operons. En aktivator binder seg i det regulatoriske området for en operon, hjelpe RNA polymerase binder seg til selskapet, og dermed øke transkripsjon av dette operon. En induser påvirkninger transkripsjon gjennom samspill med en repressor eller aktivator.
- trp operon er et klassisk eksempel på en repressible operon., Når tryptofan akkumuleres, tryptofan binder seg til en repressor, som deretter binder seg til operatør, hindre ytterligere transkripsjon.
- lac operon er et klassisk eksempel en inducible operon. Når laktose er til stede i cellen, det er konvertert til allolactose. Allolactose fungerer som en induser, binding til repressor og hindre repressor fra binding til en operatør. Dette gjør transkripsjon av strukturelle gener.
- lac operon er også gjenstand for aktivering., Når blodsukkeret er utladet, noen cellular ATP er omgjort til leiren, som binder seg til catabolite aktivator protein (CAP). CAMP-CAP komplekse aktivere transkripsjon av lac operon. Når blodsukkeret er høyt, sin tilstedeværelse hindrer transkripsjon av lac operon og andre operons av catabolite undertrykkelse.
- Små intracellulære molekyler som kalles alarmones er laget i respons til ulike miljømessige stressfaktorer, slik at bakterier til å styre transkripsjon av en gruppe av operons, kalles en regulon.,
- Bakterier har evnen til å endre hvilke σ faktor av RNA polymerase de bruker i respons til miljømessige forhold til å raskt og globalt endring som regulons er transkribert.
- Prokaryotes har regulatoriske mekanismer, inkludert demping og bruk av riboswitches, for samtidig å styre gjennomføring av transkripsjon og oversettelse fra at transkripsjonen. Disse mekanismene fungerer gjennom dannelsen av stammen looper i 5′ – enden av en mRNA-molekylet tiden blir syntetisert.,
- Det er ekstra poeng for regulering av genuttrykk i prokaryotes og eukaryotes. I eukaryotes, epigenetisk regulering av kjemisk modifikasjon av DNA eller histones, og regulering av RNA-prosessering er to metoder.
Flere Valg
En operon av genene som koder for enzymer i en biosynthetic vei er sannsynlig å være hvilke av de følgende?
- inducible
- repressible
- første
- monocistronic
En operon koding gener som er transkribert og oversatt kontinuerlig for å gi cellen med konstant mellomliggende nivåer av protein produkter sies å være noe av følgende?
- repressible
- inducible
- første
- aktivert
Hvilke av følgende forhold fører til maksimal uttrykk av lac operon?
- laktose til stede, glukose fraværende
- laktose til stede, glukose presentere
- laktose fraværende, glukose fraværende
- laktose fraværende, glukose presentere
Hvilken av de følgende er en type regulering av genuttrykk som er unike for eukaryotes?,
- demping
- bruk av alternative σ faktor
- kjemisk modifikasjon av histones
- alarmones
Fyll ut Blank
Den DNA-sekvens, som repressors kan binde, som ligger mellom arrangøren og den første strukturelle genet kalles ________.,
forebygging av uttrykk av operons koding substrat bruk trasé for andre underlag enn glukose når glukose er til stede kalles _______.
Tenk på Det
- Det er to måter som bakterier kan påvirke transkripsjon av flere forskjellige operons samtidig i respons til et bestemt miljø-tilstand?
- følgende figur er fra Monod opprinnelige arbeid på diauxic vekst som viser vekst av E. coli i samtidig tilstedeværelse av xylose og glukose som bare karbon kilder. Forklare hva som skjer under punktene A–D med hensyn til carbon kilde som er brukt for vekst, og forklare om xylose-bruk operon blir uttrykt (og hvorfor)., Vær oppmerksom på at uttrykk av de enzymene som er nødvendige for xylose bruk er regulert på en måte som ligner til uttrykk i de enzymene som er nødvendige for laktose bruk.
