Lernziele
- Vergleichen Sie induzierbare Operonen und repressible Operonen
- Beschreiben Sie, warum die Regulation von Operonen wichtig ist
Jede Kernzelle in einem vielzelligen Organismus enthält Kopien derselben DNA. In ähnlicher Weise enthalten alle Zellen in zwei reinen Bakterienkulturen, die aus derselben Ausgangskolonie geimpft wurden, dieselbe DNA, mit Ausnahme von Veränderungen, die durch spontane Mutationen entstehen., Wenn jede Zelle in einem vielzelligen Organismus die gleiche DNA hat, wie können dann Zellen in verschiedenen Körperteilen des Organismus unterschiedliche Eigenschaften aufweisen? Wie kommt es, dass dieselben Bakterienzellen in zwei Reinkulturen, die unterschiedlichen Umweltbedingungen ausgesetzt sind, unterschiedliche Phänotypen aufweisen können? In beiden Fällen aktiviert oder exprimiert jede genetisch identische Zelle nicht denselben Satz von Genen. Es wird nur eine Teilmenge von Proteinen in einer Zelle zu einem bestimmten Zeitpunkt exprimiert.,
Genomische DNA enthält sowohl Strukturgene, die Produkte kodieren, die als zelluläre Strukturen oder Enzyme dienen, als auch regulatorische Gene, die Produkte kodieren, die die Genexpression regulieren. Die Expression eines Gens ist ein stark regulierter Prozess. Während die Regulierung der Genexpression in mehrzelligen Organismen eine zelluläre Differenzierung ermöglicht, wird in einzelligen Organismen wie Prokaryoten in erster Linie sichergestellt, dass die Ressourcen einer Zelle nicht verschwendet werden, um Proteine herzustellen, die die Zelle zu diesem Zeitpunkt nicht benötigt.,
Die Aufklärung der Mechanismen, die die Genexpression steuern, ist wichtig für das Verständnis der menschlichen Gesundheit. Fehlfunktionen in diesem Prozess beim Menschen führen zur Entwicklung von Krebs und anderen Krankheiten. Das Verständnis der Wechselwirkung zwischen der Genexpression eines Erregers und der seines menschlichen Wirts ist wichtig für das Verständnis einer bestimmten Infektionskrankheit. Die Genregulation beinhaltet ein komplexes Netz von Interaktionen innerhalb einer bestimmten Zelle zwischen Signalen aus der Zellumgebung, Signalmolekülen innerhalb der Zelle und der DNA der Zelle., Diese Wechselwirkungen führen je nach Umständen zur Expression einiger Gene und zur Unterdrückung anderer.
Prokaryoten und Eukaryoten haben einige Ähnlichkeiten in ihren Mechanismen zur Regulierung der Genexpression; Die Genexpression bei Eukaryoten ist jedoch aufgrund der zeitlichen und räumlichen Trennung zwischen den Prozessen der Transkription und Translation komplizierter., Obwohl die meiste Regulation der Genexpression durch Transkriptionskontrolle bei Prokaryoten erfolgt, erfolgt die Regulation der Genexpression bei Eukaryoten auf Transkriptionsebene und posttranskriptional (nachdem das primäre Transkript erstellt wurde).
Prokaryotische Genregulation
In Bakterien und Archaeen werden normalerweise Strukturproteine mit verwandten Funktionen innerhalb des Genoms in einem als Operon bezeichneten Block codiert und zusammen unter der Kontrolle eines einzelnen Promotors transkribiert, was zur Bildung eines polyzistronen Transkripts führt (Abbildung 1)., Auf diese Weise kann die Regulation der Transkription aller Strukturgene, die für die Enzyme kodieren, die die vielen Schritte in einem einzigen biochemischen Weg katalysieren, gleichzeitig gesteuert werden, da sie entweder alle gleichzeitig benötigt werden oder keine benötigt wird. In E. coli liegen beispielsweise alle Strukturgene, die Enzyme kodieren, die zur Verwendung von Lactose als Energiequelle erforderlich sind, nebeneinander im Lactose (oder lac) – Operon unter der Kontrolle eines einzelnen Promotors, des Lac-Promotors., Die französischen Wissenschaftler François Jacob (1920-2013) und Jacques Monod am Pasteur-Institut zeigten als erste die Organisation bakterieller Gene in Operonen durch ihre Studien zum Lac Operon von E. coli. Für diese Arbeit gewannen sie 1965 den Nobelpreis für Physiologie oder Medizin. Obwohl eukaryotische Gene nicht in Operonen organisiert sind, sind prokaryotische Operonen hervorragende Modelle, um allgemein etwas über die Genregulation zu lernen. Es gibt einige Gencluster in Eukaryoten, die ähnlich wie Operonen funktionieren., Viele der Prinzipien können auf eukaryotische Systeme angewendet werden und tragen zu unserem Verständnis von Veränderungen der Genexpression in Eukaryoten bei, die zu pathologischen Veränderungen wie Krebs führen können.

Abbildung 1. Bei Prokaryoten werden Strukturgene verwandter Funktionen häufig zusammen auf dem Genom organisiert und zusammen unter der Kontrolle eines einzelnen Promotors transkribiert. Der Regulierungsbereich von operon umfasst sowohl den Projektträger als auch den Betreiber. Wenn ein Repressor an den Operator bindet, werden die Strukturgene nicht transkribiert., Alternativ können Aktivatoren an den regulatorischen Bereich binden und die Transkription verbessern.
Jedes Operon enthält DNA-Sequenzen, die seine eigene Transkription beeinflussen; Diese befinden sich in einer Region, die als regulatorische Region bezeichnet wird. Der regulatorische Bereich umfasst den Promotor und den den Promotor umgebenden Bereich, an den Transkriptionsfaktoren, Proteine, die durch regulatorische Gene kodiert werden, binden können. Transkriptionsfaktoren beeinflussen die Bindung von RNA-Polymerase an den Promotor und ermöglichen dessen Fortschreiten, um Strukturgene zu transkribieren., Ein Repressor ist ein Transkriptionsfaktor, der die Transkription eines Gens als Reaktion auf einen externen Stimulus unterdrückt, indem er an eine DNA-Sequenz innerhalb des regulatorischen Bereichs namens Operator bindet, der sich zwischen der RNA-Polymerase-Bindungsstelle des Promotors und der Transkriptionsstartstelle des ersten Strukturgens befindet. Die Repressorbindung blockiert physikalisch die RNA-Polymerase von der Transkription struktureller Gene., Umgekehrt ist ein Aktivator ein Transkriptionsfaktor, der die Transkription eines Gens als Reaktion auf einen externen Stimulus erhöht, indem er die RNA-Polymerase-Bindung an den Promotor erleichtert. Ein Induktor, ein dritter Typ von Regulationsmolekül, ist ein kleines Molekül, das entweder die Transkription aktiviert oder unterdrückt, indem es mit einem Repressor oder einem Aktivator interagiert.
In Prokaryoten gibt es Beispiele für Operonen, deren Genprodukte ziemlich konsistent benötigt werden und deren Expression daher unreguliert ist., Solche Operonen werden konstitutiv exprimiert, was bedeutet, dass sie kontinuierlich transkribiert und übersetzt werden, um die Zelle mit konstanten Zwischenspiegeln der Proteinprodukte zu versorgen. Solche Gene kodieren Enzyme, die an den Funktionen beteiligt sind, die für die zelluläre Aufrechterhaltung erforderlich sind, einschließlich DNA-Replikation, – reparatur und-expression, sowie Enzyme, die am Kernstoffwechsel beteiligt sind. Im Gegensatz dazu gibt es andere prokaryotische Operonen, die nur bei Bedarf exprimiert werden und durch Repressoren, Aktivatoren und Induktoren reguliert werden.,
Denken Sie darüber nach
- Was sind die Teile in der DNA-Sequenz eines Operons?
- Welche Arten von regulatorischen Molekülen gibt es?
Regulation durch Repression
Prokaryotische Operonen werden üblicherweise durch die Bindung von Repressoren an Operatorregionen gesteuert, wodurch die Transkription der Strukturgene verhindert wird. Solche Operonen werden entweder als repressible Operonen oder induzierbare Operonen klassifiziert. Repressible Operonen, wie das Tryptophan (trp) – Operon, enthalten typischerweise Gene, die Enzyme kodieren, die für einen biosynthetischen Weg benötigt werden., Solange das Produkt des Weges, wie Tryptophan, weiterhin von der Zelle benötigt wird, wird weiterhin ein repressibles Operon exprimiert. Wenn sich jedoch das Produkt des Biosyntheseweges in der Zelle ansammelt und die Notwendigkeit beseitigt, dass die Zelle weiterhin mehr bildet, wird die Expression des Operons unterdrückt. Umgekehrt enthalten induzierbare Operonen, wie das Lac-Operon von E. coli, oft Gene, die Enzyme in einem Weg kodieren, der am Metabolismus eines bestimmten Substrats wie Laktose beteiligt ist., Diese Enzyme werden nur benötigt, wenn dieses Substrat verfügbar ist, so dass die Expression der Operonen typischerweise nur in Gegenwart des Substrats induziert wird.
Das Trp-Operon: Ein repressibles Operon
E. coli kann Tryptophan unter Verwendung von Enzymen synthetisieren, die von fünf Strukturgenen codiert werden, die sich nebeneinander im Trp-Operon befinden (Abbildung 2). Wenn das Tryptophan niedrig ist, wird das Operon eingeschaltet. Dies bedeutet, dass die Transkription initiiert, die Gene exprimiert und Tryptophan synthetisiert wird., Wenn jedoch Tryptophan in der Umgebung vorhanden ist, wird das TRP-Operon ausgeschaltet. Transkription tritt nicht auf und Tryptophan wird nicht synthetisiert.
Wenn Tryptophan nicht in der Zelle vorhanden ist, bindet der Repressor selbst nicht an den Operator; Daher ist das Operon aktiv und Tryptophan wird synthetisiert. Wenn sich jedoch Tryptophan in der Zelle ansammelt, binden zwei Tryptophanmoleküle an das Trp-Repressormolekül, das seine Form ändert und es ihm ermöglicht, sich an den TRP-Operator zu binden., Diese Bindung der aktiven Form des trp-Repressors an den Operator blockiert die RNA-Polymerase daran, die Strukturgene zu transkribieren, wodurch die Expression des Operons gestoppt wird. Somit reguliert das tatsächliche Produkt des vom Operon gesteuerten Biosyntheseweges die Expression des Operons.
Abbildung 2. Die fünf Strukturgene, die zur Synthese von Tryptophan in E. coli benötigt werden, befinden sich im Trp-Operon nebeneinander. Wenn Tryptophan fehlt, bindet das Repressorprotein nicht an den Operator und die Gene werden transkribiert., Wenn Tryptophan reichlich vorhanden ist, bindet Tryptophan das Repressorprotein an der Operatorsequenz. Dies blockiert physikalisch die RNA-Polymerase von der Transkription der Tryptophan-Biosynthese-Gene.
Sehen Sie sich dieses Video an, um mehr über das trp-Operon zu erfahren.
Das Lac-Operon: Ein induzierbares Operon
Das Lac-Operon ist ein Beispiel für ein induzierbares Operon, das auch in Abwesenheit von Glukose aktiviert wird (Abbildung 3)., Das Lac Operon kodiert drei Strukturgene, die notwendig sind, um das Disaccharid Lactose aus der Umwelt zu gewinnen und zu verarbeiten, und zerlegt es in die einfachen Zucker Glucose und Galactose. Damit das Lakridon exprimiert werden kann, muss Laktose vorhanden sein. Dies ist für die Zelle sinnvoll, da es energetisch verschwenderisch wäre, die Enzyme zur Verarbeitung von Laktose zu erzeugen, wenn keine Laktose verfügbar wäre.
In Abwesenheit von Laktose ist der Lac-Repressor an die Operatorregion des Lac-Operons gebunden, wodurch physikalisch verhindert wird, dass die RNA-Polymerase die Strukturgene transkribiert., Wenn jedoch Laktose vorhanden ist, wird die Laktose in der Zelle in Allolactose umgewandelt. Allolactose dient als Induktormolekül, das an den Repressor bindet und seine Form ändert, so dass es nicht mehr an die Operator-DNA binden kann. Die Entfernung des Repressors in Gegenwart von Lactose ermöglicht es der RNA-Polymerase, sich durch die Operatorregion zu bewegen und mit der Transkription der LAC-Strukturgene zu beginnen.
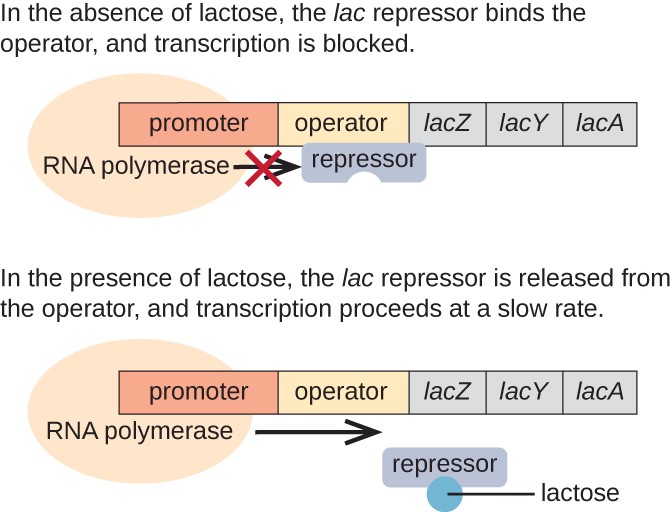
Abbildung 3. Die drei Strukturgene, die zum Abbau von Laktose in E. coli benötigt werden, befinden sich nebeneinander im Lac Operon., Wenn Laktose fehlt, bindet das Repressorprotein an den Operator, wodurch die RNA-Polymerase physisch daran gehindert wird, die LAC-Strukturgene zu transkribieren. Wenn Laktose verfügbar ist, bindet ein Laktosemolekül das Repressorprotein, wodurch verhindert wird, dass der Repressor an die Operatorsequenz bindet, und die Gene werden transkribiert.
Der lac-Operon: Aktivierung durch Katabolitenaktivatorprotein

Abbildung 4. Wenn E. coli in Gegenwart von zwei Substraten gezüchtet wird, verwendet es das bevorzugte Substrat (in diesem Fall Glukose), bis es erschöpft ist., Dann werden Enzyme, die für den Metabolismus des zweiten Substrats benötigt werden, exprimiert und das Wachstum wird wieder aufgenommen, wenn auch langsamer.
Bakterien haben typischerweise die Fähigkeit, eine Vielzahl von Substraten als Kohlenstoffquellen zu verwenden. Da Glukose jedoch normalerweise anderen Substraten vorzuziehen ist, haben Bakterien Mechanismen, um sicherzustellen, dass alternative Substrate nur verwendet werden, wenn Glukose aufgebraucht ist. Darüber hinaus verfügen Bakterien über Mechanismen, um sicherzustellen, dass die Gene, die Enzyme für die Verwendung alternativer Substrate kodieren, nur exprimiert werden, wenn das alternative Substrat verfügbar ist., In den 1940er Jahren demonstrierte Jacques Monod als erster die Präferenz für bestimmte Substrate gegenüber anderen durch seine Studien über das Wachstum von E. coli, wenn es in Gegenwart von zwei verschiedenen Substraten gleichzeitig kultiviert wurde. Solche Studien erzeugten diauxische Wachstumskurven, wie die in Abbildung 4 gezeigte. Obwohl zuerst das bevorzugte Substrat Glucose verwendet wird, wächst E. coli schnell und die Enzyme für den Laktosestoffwechsel fehlen. Sobald der Glukosespiegel jedoch aufgebraucht ist, verlangsamen sich die Wachstumsraten und induzieren die Expression der Enzyme, die für den Stoffwechsel des zweiten Substrats, der Laktose, benötigt werden., Beachten Sie, wie die Wachstumsrate in Laktose langsamer ist, wie durch die geringere Steilheit der Wachstumskurve angezeigt.
Die Fähigkeit, von der Glukoseverwendung zu einem anderen Substrat wie Laktose zu wechseln, ist eine Folge der Aktivität eines Enzyms namens Enzym IIA (EIIA). Wenn der Glukosespiegel sinkt, produzieren die Zellen weniger ATP aus dem Katabolismus (siehe Katabolismus von Kohlenhydraten), und EIIA wird phosphoryliert., Phosphoryliertes EIIA aktiviert Adenylylcyclase, ein Enzym, das einen Teil des verbleibenden ATP in cyclisches AMP (cAMP) umwandelt, ein zyklisches Derivat von AMP und wichtiges Signalmolekül, das am Glukose-und Energiestoffwechsel in E. coli beteiligt ist. Infolgedessen beginnen die cAMP-Werte in der Zelle zu steigen (Abbildung 5).

Abbildung 5. Wenn der ATP-Spiegel aufgrund des Glukoseabbaus abnimmt, wird ein Teil des verbleibenden ATP durch Adenylylcyclase in cAMP umgewandelt. Daher signalisieren erhöhte cAMP-Spiegel einen Glukoseabbau.,
Auch bei der Umstellung von Glukose auf Laktose spielt der Lactose eine Rolle. Wenn Glukose knapp ist, bindet das akkumulierende cAMP, das durch erhöhte Adenylylcyclaseaktivität verursacht wird, an das Katabolitenaktivatorprotein (CAP), das auch als cAMP-Rezeptorprotein (CRP) bekannt ist. Der Komplex bindet an die Promotorregion des Lac Operons (Abbildung 6). In den regulatorischen Regionen dieser Operonen befindet sich eine Kappenbindungsstelle stromaufwärts der RNA-Polymerase-Bindungsstelle im Promotor., Die Bindung des CAP-cAMP-Komplexes an diese Stelle erhöht die Bindungsfähigkeit der RNA-Polymerase an die Promotorregion, um die Transkription der Strukturgene zu initiieren. Im Falle des Lac-Operons muss daher für die Transkription Laktose vorhanden sein (Entfernung des Lac-Repressorproteins) und der Glukosespiegel muss aufgebraucht sein (Bindung eines aktivierenden Proteins ermöglichen). Wenn der Glukosespiegel hoch ist, kommt es zur Katabolitenrepression von Operonen, die Enzyme für den Stoffwechsel alternativer Substrate kodieren., Aufgrund des niedrigen cAMP-Levels unter diesen Bedingungen gibt es eine unzureichende Menge des CAP-cAMP-Komplexes, um die Transkription dieser Operonen zu aktivieren. Siehe Tabelle 1 für eine Zusammenfassung der Regelung des lac Operon.

Abbildung 6. (a) In Gegenwart von cAMP bindet CAP an die Promotoren von Operonen, wie das Lac-Operon, die Gene für Enzyme für die Verwendung alternativer Substrate kodieren. (b) Damit der Lac-Operon ausgedrückt werden kann, muss eine Aktivierung durch cAMP-CAP sowie die Entfernung des Lac-Repressors vom Bediener erfolgen.,
| Tabelle 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
Denken Sie darüber nach
- Was beeinflusst die Bindung des trp Operon Repressors an den Operator?
- Wie und wann wird das Verhalten des lac repressor Proteins verändert?
- Wie sonst ist der lac operon nicht nur repressibel, sondern auch regulierbar?
Globale Reaktionen von Prokaryoten
Bei Prokaryoten gibt es auch mehrere höhere Ebenen der Genregulation, die die Transkription vieler verwandter Operonen gleichzeitig als Reaktion auf ein Umweltsignal steuern können., Eine Gruppe von Operonen, die alle gleichzeitig gesteuert werden, wird Regulon genannt.
Alarmone
Wenn Sie drohenden Stress spüren, verändern Prokaryoten die Expression einer Vielzahl von Operonen, um koordiniert zu reagieren. Sie tun dies durch die Produktion von Alarmonen, die kleine intrazelluläre Nukleotidderivate sind. Alarmone verändern, welche Gene exprimiert werden und stimulieren die Expression spezifischer Stressreaktionsgene. Die Verwendung von Alarmonen zur Veränderung der Genexpression als Reaktion auf Stress scheint bei pathogenen Bakterien wichtig zu sein., Bei der Begegnung mit Wirtsabwehrmechanismen und anderen rauen Bedingungen während der Infektion werden viele Operonen, die für Virulenzgene kodieren, als Reaktion auf die Alarmonsignalisierung hochreguliert. Die Kenntnis dieser Reaktionen ist der Schlüssel, um den Infektionsprozess vieler Krankheitserreger vollständig zu verstehen und Therapien zu entwickeln, um diesem Prozess entgegenzuwirken.,
Alternative σ-Faktoren
Da die σ-Untereinheit der bakteriellen RNA-Polymerase die Spezifität verleiht, welche Promotoren transkribiert werden sollen, ist die Änderung des verwendeten σ-Faktors eine weitere Möglichkeit für Bakterien, schnell und global zu ändern, welche Regulonen zu einem bestimmten Zeitpunkt transkribiert werden. Der σ-Faktor erkennt Sequenzen innerhalb eines bakteriellen Promotors, so dass unterschiedliche σ-Faktoren jeweils leicht unterschiedliche Promotorsequenzen erkennen., Auf diese Weise kann die Zelle, wenn sie bestimmte Umweltbedingungen wahrnimmt, reagieren, indem sie ändert, welchen σ-Faktor sie exprimiert, den alten abbaut und einen neuen erzeugt, um die Operonen zu transkribieren, die für Gene kodieren, deren Produkte unter dem neuen Umgebungszustand nützlich sind. Zum Beispiel steuert bei sporulierenden Bakterien der Gattungen Bacillus und Clostridium (zu denen viele Krankheitserreger gehören) eine Gruppe von genetischen Faktoren die Expression der vielen Gene, die für die Sporulation als Reaktion auf sporulationsstimulierende Signale benötigt werden.,
Denken Sie darüber nach
- Wie heißt eine Sammlung von Operonen, die als Gruppe reguliert werden können?
- Welche Art von Reiz würde die Transkription eines anderen σ-Faktors auslösen?
Zusätzliche Regulierungsmethoden bei Bakterien: Dämpfung und Riboschalter
Obwohl die meisten Genexpressionen auf der Ebene der Transkriptionsinitiation bei Prokaryoten reguliert werden, gibt es auch Mechanismen, um sowohl den Abschluss der Transkription als auch die gleichzeitige Translation zu kontrollieren., Seit ihrer Entdeckung wurde gezeigt, dass diese Mechanismen den Abschluss der Transkription und Übersetzung vieler prokaryotischer Operonen steuern. Da diese Mechanismen die Regulierung von Transkription und Übersetzung direkt miteinander verbinden, sind sie spezifisch für Prokaryoten, da diese Prozesse in Eukaryoten physisch getrennt sind.
Ein solches Regelungssystem ist die Dämpfung, wobei sekundäre Stammschleifenstrukturen, die innerhalb des 5′ – Endes einer transkribierten mRNA gebildet werden, bestimmen, ob eine Transkription zur Vervollständigung der Synthese dieser mRNA stattfindet und ob diese mRNA für die Translation verwendet wird., Über den bereits diskutierten transkriptionellen Repressionsmechanismus hinaus steuert die Dämpfung auch die Expression des trp-Operons in E. coli (Abbildung 7). Der trp-Operon-regulatorische Bereich enthält eine führende Sequenz namens trpL zwischen dem Operator und dem ersten Strukturgen, die vier RNA-Abschnitte aufweist, die in verschiedenen Kombinationen miteinander basenpaarweise sein können. Wenn sich eine Terminatorstammschleife bildet, endet die Transkription und setzt RNA-Polymerase aus der mRNA frei., Wenn sich jedoch eine Antiterminator-Stammschleife bildet, verhindert dies die Bildung der Terminator-Stammschleife, so dass RNA-Polymerase die Strukturgene transkribieren kann.
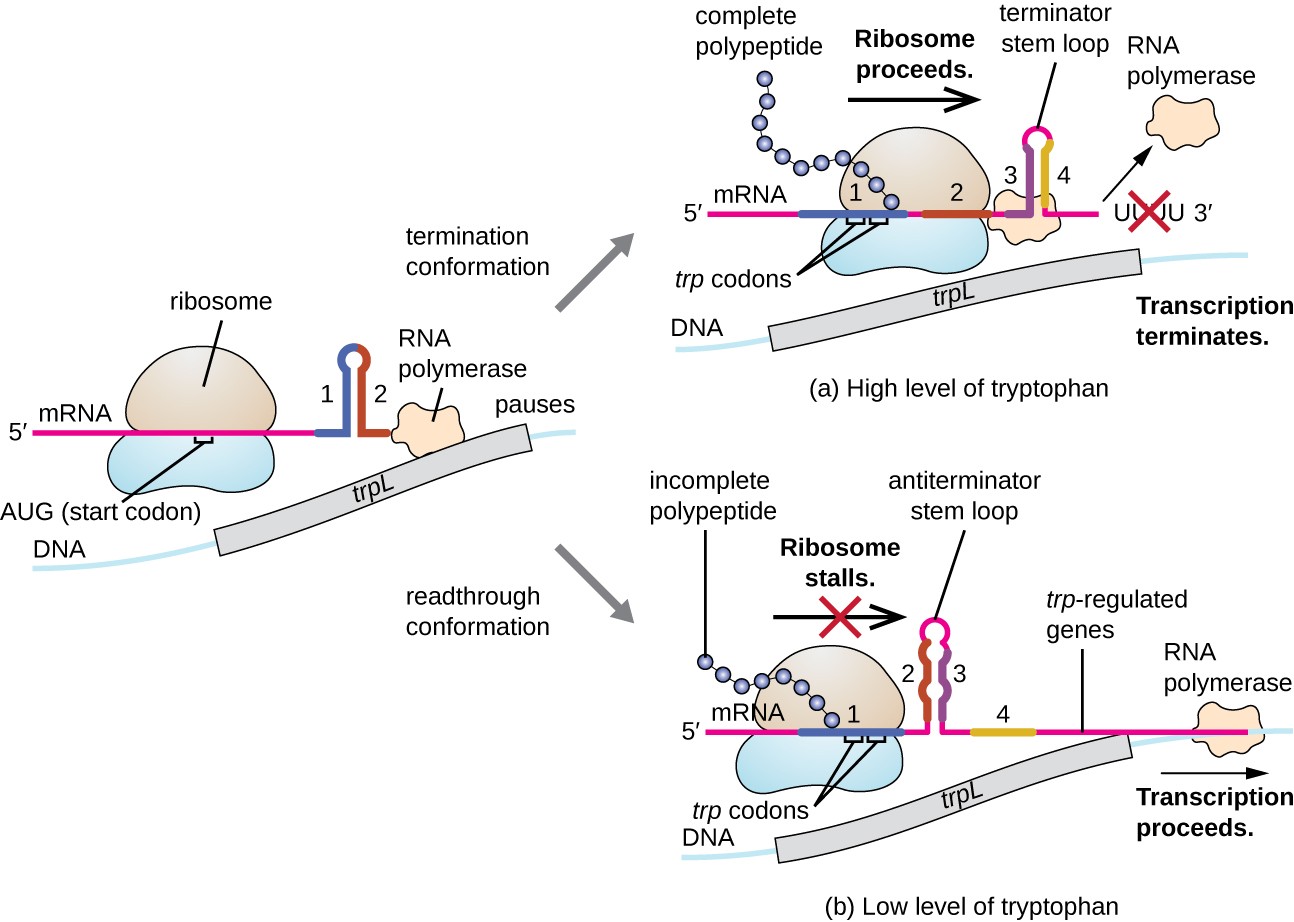
Abbildung 7. Klicken Sie, um ein größeres Bild anzuzeigen. Wenn Tryptophan reichlich vorhanden ist, erfolgt die Übersetzung des durch trpL codierten Kurzzeitpeptids, die Terminatorschleife zwischen den Regionen 3 und 4 bildet sich und die Transkription endet., Wenn die Tryptophanspiegel erschöpft sind, bleibt die Translation des Short Leader-Peptids in Region 1 bestehen, sodass die Regionen 2 und 3 eine Antiterminatorschleife bilden können, und die RNA-Polymerase kann die Strukturgene des TRP-Operons transkribieren.
Ein verwandter Mechanismus der gleichzeitigen Regulation von Transkription und Translation in Prokaryoten ist die Verwendung eines Riboswitches, einer kleinen Region nicht codierender RNA, die sich am 5′ – Ende einiger prokaryotischer mRNA-Moleküle befindet (Abbildung 8). Ein Riboschalter kann an ein kleines intrazelluläres Molekül binden, um bestimmte Sekundärstrukturen des mRNA-Moleküls zu stabilisieren., Die Bindung des kleinen Moleküls bestimmt, welche Stammschleifenstruktur sich bildet, und beeinflusst so den Abschluss der mRNA-Synthese und Proteinsynthese.

Abbildung 8. Klicken Sie für ein größeres Bild. Riboswitches, die in prokaryotischen mRNA-Molekülen gefunden werden, können an kleine intrazelluläre Moleküle binden, bestimmte RNA-Strukturen stabilisieren und entweder den Abschluss der Synthese des mRNA-Moleküls selbst (links) oder das Protein beeinflussen, das mit dieser mRNA hergestellt wurde (rechts).,
Andere Faktoren, die die Genexpression in Prokaryoten und Eukaryoten beeinflussen
Obwohl der Fokus auf unserer Diskussion der Transkriptionskontrolle prokaryotische Operonen als Beispiele verwendete, ist die eukaryotische Transkriptionskontrolle in vielerlei Hinsicht ähnlich. Wie bei Prokaryoten kann die eukaryotische Transkription durch die Bindung von Transkriptionsfaktoren einschließlich Repressoren und Aktivatoren gesteuert werden., Interessanterweise kann die eukaryotische Transkription durch die Bindung von Proteinen an DNA-Regionen, sogenannte Enhancer, ziemlich weit vom Gen entfernt, durch DNA-Schleifen beeinflusst werden, die zwischen dem Enhancer und dem Promotor erleichtert werden (Abbildung 9). Insgesamt ist die Regulierung der Transkription ein hochwirksamer Weg, um die Genexpression sowohl in Prokaryoten als auch in Eukaryoten zu kontrollieren. Die Kontrolle der Genexpression in Eukaryoten als Reaktion auf Umwelt-und zelluläre Belastungen kann jedoch auf zusätzliche Weise ohne Bindung von Transkriptionsfaktoren an regulatorische Regionen durchgeführt werden.,

Abbildung 9. In Eukaryoten ist ein Enhancer eine DNA-Sequenz, die die Transkription fördert. Jeder Enhancer besteht aus kurzen DNA-Sequenzen, die als distale Kontrollelemente bezeichnet werden. An die distalen Steuerelemente gebundene Aktivatoren interagieren mit Mediatorproteinen und Transkriptionsfaktoren. Zwei verschiedene Gene können denselben Promotor, aber unterschiedliche distale Kontrollelemente aufweisen, was eine differenzielle Genexpression ermöglicht.,
Kontrolle auf DNA-Ebene
Bei Eukaryoten können die DNA-Moleküle oder assoziierten Histone chemisch so modifiziert werden, dass sie die Transkription beeinflussen; dies wird als epigenetische Regulation bezeichnet. Es hat sich gezeigt, dass die Methylierung bestimmter Cytosin-Nukleotide in DNA als Reaktion auf Umweltfaktoren die Verwendung solcher DNA für die Transkription beeinflusst, wobei die DNA-Methylierung üblicherweise mit niedrigeren Niveaus der Genexpression korreliert., Darüber hinaus können Histonproteine für die Verpackung von DNA als Reaktion auf Umweltfaktoren auch auf vielfältige Weise chemisch modifiziert werden, einschließlich Acetylierung und Desacetylierung, was den Verpackungszustand von DNA beeinflusst und somit die Verfügbarkeit von lose gewickelter DNA für die Transkription beeinflusst. Diese chemischen Modifikationen können manchmal durch mehrere Runden der Zellteilung aufrechterhalten werden, wodurch zumindest einige dieser epigenetischen Veränderungen vererbbar werden.
Dieses Video beschreibt, wie die epigenetische Regulation die Genexpression steuert.,
Denken Sie darüber nach
- Was stoppt oder erlaubt die Transkription, wenn die Dämpfung in Betrieb ist?
- Was bestimmt den Zustand einer riboswitch?
- Beschreiben Sie die Funktion eines Enhancers.
- Beschreiben zwei Mechanismen der epigenetischen Regulation in Eukaryoten.
Klinischer Fokus: Travis, Auflösung
Dieses Beispiel schließt Travis ‚ Geschichte ab, die in den Funktionen genetisches Material, RNA-Transkription und wie asexuelle Prokaryoten genetische Vielfalt erreichen.,
Obwohl Travis seinen Kampf mit nekrotisierender Fasziitis überlebte, musste er sich nun einer Hauttransplantation unterziehen, gefolgt von einer Langzeittherapie. Aufgrund der Menge an Muskelmasse, die er verloren hat, ist es unwahrscheinlich, dass sein Bein wieder voll belastbar ist, aber sein Physiotherapeut ist optimistisch, dass er sein Bein wieder einsetzen wird.
Labortests ergaben, dass der Erreger der Travis-Infektion ein Streptokokkenstamm der Gruppe A (Strep der Gruppe A) war., Wie gesetzlich vorgeschrieben, wurde Travis ‚ Fall dem State Health Department und letztendlich den Centers for Disease Control and Prevention (CDC) gemeldet. Bei der CDC wurde der aus Travis isolierte Strep-Stamm der Gruppe A gründlicher auf Methicillinresistenz analysiert.
Methicillinresistenz ist genetisch kodiert und wird in Strep der Gruppe A durch horizontalen Gentransfer immer häufiger. Bei nekrotisierender Fasziitis ist der Blutfluss in den infizierten Bereich typischerweise aufgrund der Wirkung verschiedener genetisch kodierter bakterieller Toxine begrenzt., Aus diesem Grund kommt es aufgrund des Inzisionstests typischerweise zu wenig bis gar keiner Blutung. Leider begrenzen diese bakteriellen Toxine die Wirksamkeit intravenöser Antibiotika bei der Beseitigung von Infektionen von der Haut und dem darunter liegenden Gewebe, was bedeutet, dass die Antibiotikaresistenz allein die Unwirksamkeit der Behandlung von Travis nicht erklärt. Dennoch war eine intravenöse Antibiotikatherapie gerechtfertigt, um das mögliche Ergebnis einer Sepsis, die ein häufiges Ergebnis einer nekrotisierenden Fasziitis ist, zu minimieren., Durch die genomische Analyse des aus Travis isolierten Stammes durch die CDC wurde gezeigt, dass mehrere der wichtigen Virulenzgene auf Prophagen kodiert sind, was darauf hinweist, dass die Transduktion für den horizontalen Gentransfer dieser Gene von einer Bakterienzelle zur anderen wichtig ist.
Schlüsselkonzepte und Zusammenfassung
- Die Genexpression ist ein streng regulierter Prozess.
- Die Genexpression in Prokaryoten ist zum Zeitpunkt der Transkription weitgehend reguliert. Die Genexpression in Eukaryoten wird zusätzlich post-transkriptionell reguliert.,
- Prokaryotische Strukturgene verwandter Funktionen sind häufig in Operonen organisiert, die alle durch Transkription von einem einzelnen Promotor gesteuert werden. Der regulatorische Bereich eines Operons umfasst den Promotor selbst und den den Promotor umgebenden Bereich, an den Transkriptionsfaktoren binden können, um die Transkription zu beeinflussen.
- Obwohl einige Operonen konstitutiv ausgedrückt werden, unterliegen die meisten der Regulierung durch die Verwendung von Transkriptionsfaktoren (Repressoren und Aktivatoren)., Ein Repressor bindet an einen Operator, eine DNA-Sequenz innerhalb des regulatorischen Bereichs zwischen der RNA-Polymerase-Bindungsstelle im Promotor und dem ersten Strukturgen, wodurch die Transkription dieser Operonen physikalisch blockiert wird. Ein Aktivator bindet innerhalb des regulatorischen Bereichs eines Operons und hilft der RNA-Polymerase, sich an den Promotor zu binden, wodurch die Transkription dieses Operons verbessert wird. Ein Induktor beeinflusst die Transkription durch Interaktion mit einem repressor oder Aktivator.
- Das trp-operon, ist ein klassisches Beispiel für eine repressible operon., Wenn sich Tryptophan ansammelt, bindet Tryptophan an einen Repressor, der dann an den Operator bindet und eine weitere Transkription verhindert.
- Der lac Operon ist ein klassisches Beispiel eines induzierbaren Operons. Wenn Laktose in der Zelle vorhanden ist, wird sie in Allolactose umgewandelt. Allolactose wirkt als Induktor, bindet an den Repressor und verhindert, dass der Repressor an den Bediener bindet. Dies ermöglicht die Transkription der Strukturgene.
- Der lac Operon unterliegt ebenfalls der Aktivierung., Wenn der Glukosespiegel erschöpft ist, wird etwas zelluläres ATP in cAMP umgewandelt, das an das Katabolitenaktivatorprotein (CAP) bindet. Der cAMP-CAP-Komplex aktiviert die Transkription des lac-operon. Wenn der Glukosespiegel hoch ist, verhindert seine Anwesenheit die Transkription des Lac-Operons und anderer Operonen durch Katabolitenrepression.
- Kleine intrazelluläre Moleküle, sogenannte Alarmone, werden als Reaktion auf verschiedene Umweltbelastungen hergestellt, so dass Bakterien die Transkription einer Gruppe von Operonen, einem sogenannten Regulon, kontrollieren können.,
- Bakterien haben die Fähigkeit zu ändern, welchen σ-Faktor der RNA-Polymerase sie als Reaktion auf Umweltbedingungen verwenden, um schnell und global zu ändern, welche Regulonen transkribiert werden.
- Prokaryoten haben regulatorische Mechanismen, einschließlich Dämpfung und die Verwendung von Riboswitches, um gleichzeitig den Abschluss der Transkription und Übersetzung aus diesem Transkript zu steuern. Diese Mechanismen wirken durch die Bildung von Stammschleifen am 5′ – Ende eines mRNA-Moleküls, das derzeit synthetisiert wird.,
- Es gibt zusätzliche Punkte der Regulation der Genexpression in Prokaryoten und Eukaryoten. Bei Eukaryoten sind die epigenetische Regulation durch chemische Modifikation von DNA oder Histonen und die Regulation der RNA-Verarbeitung zwei Methoden.
Multiple Choice
Ein Operon von Genen, die Enzyme in einem Biosyntheseweg kodieren, ist wahrscheinlich welcher der folgenden?
- induzierbare
- repressible
- konstitutiven
- monocistronic
Ein Operon, das Gene kodiert, die kontinuierlich transkribiert und übersetzt werden, um die Zelle mit konstanten Zwischenspiegeln der Proteinprodukte zu versorgen, soll welches der folgenden sein?
- repressibel
- induzierbar
- konstitutiv
- aktiviert
Welche der folgenden Bedingungen führt zum maximalen Ausdruck des lac Operons?
- Laktose vorhanden, Glukose abwesend
- Laktose vorhanden, Glukose vorhanden
- Laktose abwesend, Glukose abwesend
- Laktose abwesend, Glukose vorhanden
Welche der folgenden Arten ist eine für Eukaryoten einzigartige Art der Regulation der Genexpression?,
- Abschwächung
- Verwendung eines alternativen σ-Faktors
- chemische Modifikation von Histonen
- alarmones
Füllen Sie das Leerzeichen
Die DNA-Sequenz, an die Repressoren binden können, die zwischen dem Promotor und dem ersten Strukturgen liegt, wird als ____ _ _ _ _ bezeichnet.,
Die Verhinderung der Expression von Operonen, die das Substrat codieren, verwendet Wege für andere Substrate als Glukose, wenn Glukose vorhanden ist, heißt_______.
Denken Sie darüber nach
- Was sind zwei Möglichkeiten, wie Bakterien die Transkription mehrerer verschiedener Operonen gleichzeitig als Reaktion auf eine bestimmte Umgebungsbedingung beeinflussen können?
- Die folgende Abbildung stammt aus Monods ursprünglicher Arbeit über das diauxische Wachstum, die das Wachstum von E. coli bei gleichzeitiger Anwesenheit von Xylose und Glucose als einzigen Kohlenstoffquellen zeigt. Erklären Sie, was an den Punkten A–D in Bezug auf die Kohlenstoffquelle geschieht, die für das Wachstum verwendet wird, und erklären Sie, ob das Xylose-Use-Operon ausgedrückt wird (und warum)., Beachten Sie, dass die Expression der für die Verwendung von Xylose erforderlichen Enzyme in ähnlicher Weise reguliert wird wie die Expression der für die Verwendung von Laktose erforderlichen Enzyme.
