Oppimisen Tavoitteet
- Vertaa indusoituva operons ja repressible operons
- Kuvaile miksi sääntelyä operons on tärkeää
Jokainen tumallisiin solu monisoluisen organismin sisältää kopioita samasta DNA: ta. Samoin kaikki solut kaksi puhdasta bakteeri kulttuureissa ympätty samasta alkaen siirtomaa sisältää samaa DNA: ta, lukuun ottamatta muutoksia, jotka johtuvat spontaaneja mutaatioita., Jos monisoluisessa organismissa jokaisella solulla on sama DNA, niin miten on mahdollista, että organismin eri osissa olevilla soluilla on erilaisia ominaisuuksia? Samoin, miten on mahdollista, että sama bakteeri solujen sisällä kaksi puhdasta kulttuurit altistuvat eri ympäristöolosuhteissa voi esiintyä eri fenotyypit? Kummassakaan tapauksessa jokainen geneettisesti identtinen solu ei käynnistä tai ilmaise samoja geenejä. Vain osajoukko proteiineja solussa tiettynä aikana ilmaistaan.,
Genomista DNA sisältää sekä rakenteellisia geenejä, jotka koodaavat tuotteita, jotka toimivat solujen rakenteita tai entsyymejä, ja sääntelyn geenien, jotka koodaavat tuotteita, jotka säätelevät geenien ilmentyminen. Geenin ilmentyminen on hyvin säännelty prosessi. Ottaa huomioon, että säännellä geenien ilmentyminen vuonna monisoluisten organismien avulla solujen erilaistumista, vuonna yksisoluiset organismit, kuten prokaryootit, se ensisijaisesti varmistaa, että solu on resursseja ei tuhlata tehdä proteiineja, jotka solun ei tarvitse tuolloin.,
geeniekspressiota säätelevien mekanismien selvittäminen on tärkeää ihmisten terveyden ymmärtämisen kannalta. Toimintahäiriöt tässä prosessissa ihmisillä johtavat syövän ja muiden sairauksien kehittymiseen. Taudinaiheuttajan geeniekspression ja sen ihmisisännän välisen vuorovaikutuksen ymmärtäminen on tärkeää tietyn tartuntataudin ymmärtämisen kannalta. Gene asetus edellyttää monimutkaista web vuorovaikutusta tietyn solun keskuudessa signaaleja solun ympäristöön, molekyylejä solun sisällä ja solujen DNA: ta., Nämä vuorovaikutukset johtavat joidenkin geenien ilmentymiseen ja toisten tukahduttamiseen olosuhteista riippuen.
Prokaryooteissa ja eukaryooteissa jakaa joitakin yhtäläisyyksiä niiden mekanismeja, jotka säätelevät geenien ilmentyminen; kuitenkin, geenien ilmentymisen eukariooteissa on monimutkaisempi, koska ajallinen ja paikallinen erottaminen toisistaan prosessit, transkriptio ja käännös., Näin ollen, vaikka useimmat sääntelyn geenien ilmentyminen tapahtuu kautta transkription ohjaus prokaryootit, asetuksen geenien ilmentymisen eukaryooteissa tapahtuu transkription tasolla ja post-transcriptionally (sen jälkeen, kun ensisijainen transkriptio on tehty).
Prokaryootti-Geenin Asetukseksi
bakteerien ja arkkien, rakenteellisia proteiineja liittyvät toiminnot ovat yleensä koodattu yhdessä sisällä genomin lohko kutsutaan operon ja ne litteroitiin yhdessä valvonnassa yhden toteuttajan, jolloin muodostumista polycistronic transkriptio (Kuva 1)., Tällä tavoin asetuksen transkriptio kaikki rakenteelliset geenit koodaavat entsyymejä, jotka katalysoivat monet vaiheet yksi biokemiallinen reitti voidaan ohjata samanaikaisesti, koska ne on joko kaikki tarvitaan samaan aikaan, tai ei mitään tarvitaan. Esimerkiksi E. coli, kaikki rakenteelliset geenien, jotka koodaavat entsyymejä tarvitaan käyttää laktoosia energialähteenä valehdella vierekkäin laktoosi (tai lac) operon valvonnassa yksi promoottori, lac promoottori., Ranskan tutkijat François Jacob (1920-2013) ja Jacques Monod klo Pasteur-Instituutissa olivat ensimmäinen osoittaa organisaation bakteerien geenejä operons, kautta tutkimuksissa lac-operonin E. coli. Tästä työstä he saivat Nobelin fysiologian tai lääketieteen palkinnon vuonna 1965. Vaikka aitotumallisilla geenit eivät ole jaettu operons, prokaryooteilla operons ovat erinomaisia malleja oppia gene asetus yleisesti. Eukaryooteissa on joitakin geeniklustereita, jotka toimivat samalla tavalla kuin operonit., Monet periaatteet voidaan soveltaa aitotumallisilla järjestelmiä ja edistää ymmärrystämme muutoksia geenien ilmentymisen eukariooteissa, jotka voivat aiheuttaa patologisia muutoksia, kuten syöpä.

Kuva 1. Prokaryooteissa toisiinsa liittyvien funktioiden rakenteelliset geenit organisoidaan usein yhdessä genomiin ja transkriboidaan yhdessä yhden promoottorin valvonnassa. Operonin sääntelyalueeseen kuuluvat sekä järjestäjä että operaattori. Jos repressori sitoutuu operaattoriin, rakenteellisia geenejä ei transkriboida., Vaihtoehtoisesti aktivaattorit voivat sitoutua sääntelyalueeseen tehostaen transkriptiota.
Jokainen operon sisältää DNA-sekvenssit, jotka vaikuttavat oman transkriptio; nämä sijaitsevat alueella, jota kutsutaan sääntelyn alueella. Sääntely-alue sisältää promoottori ja ympäröivällä alueella promoottori, jonka transkriptio tekijät, proteiineja koodaa sääntelyn geenejä, voi sitoa. Transkriptiotekijät vaikuttavat RNA-polymeraasin sitoutumiseen promoottoriin ja mahdollistavat sen etenemisen rakenteellisten geenien transkriptioon., A repressori on transkriptio tekijä, joka tukahduttaa transkriptio geeni vastauksena ulkoiseen ärsykkeeseen sitoutumalla DNA-sekvenssi, sääntelyn alue nimeltään operaattori, joka sijaitsee välillä RNA-polymeraasin sitoutumiskohta promoottori ja transkription alku-sivuston ensimmäisen rakenteellisen geenin. Repressorisidos estää fyysisesti RNA-polymeraasia rakenteellisten geenien transkriboinnista., Päinvastoin, aktivaattori on transkriptio tekijä, joka lisää transkriptio geeni vastauksena ulkoiseen ärsykkeeseen helpottamalla RNA-polymeraasin sitoutumisen edistäjänä. Induktori, kolmas tyyppi sääntely-molekyyli, on pieni molekyyli, joka joko aktivoi tai tukahduttaa transkriptio vuorovaikutuksessa repressori tai aktivaattori.
prokaryootit, on olemassa esimerkkejä operons, joiden geenin tuotteita tarvitaan melko johdonmukaisesti ja joiden ilme, siksi on sääntelemätöntä., Tällainen operons ovat konstitutiivisesti, eli ne kirjoitetaan puhtaaksi ja käännetään jatkuvasti tarjota solun jatkuva keskitason tasoilla proteiini tuotteita. Tällaiset geenit koodaavat entsyymejä, mukana niiden toiminnot tarvitaan solujen huolto, mukaan lukien DNA: n replikaatio -, korjaus -, ja ilmaisu, sekä osallistuvien entsyymien core aineenvaihduntaa. Sen sijaan on olemassa muita prokaryoottisia operoneja, jotka ilmaistaan vain tarvittaessa ja joita säätelevät repressorit, aktivaattorit ja induktorit.,
ajattele sitä
- mitkä ovat operonin DNA-sekvenssin osat?
- millaisia säätelymolekyylejä on olemassa?
– Asetuksen Sorron
Prokaryoottiset operons ovat yleisesti määräysvallassa sitova repressors operaattorin alueilla, mikä estää transkriptio rakenteelliset geenit. Tällaiset operonit luokitellaan joko repressiivisiksi operoneiksi tai induktiivisiksi operoneiksi. Repressiiviset operonit, kuten tryptofaani (trp) – operoni, sisältävät tyypillisesti geenejä, jotka koodaavat biosynteettiselle reitille tarvittavia entsyymejä., Niin kauan kuin tuote-reitin, kuten tryptofaania, edelleen vaaditaan solu, repressible operon jatkossakin ilmaista. Kuitenkin, kun tuote biosynteettiset koulutusjakso alkaa kerääntyä soluun, poistamalla tarve solu jatkaa tehdä enemmän, ilmaus operon on tukahdutettu. Päinvastoin, indusoituva operons, kuten lac-operonin E. coli, sisältävät usein geenit koodaavat entsyymejä väylän mukana aineenvaihduntaa tiettyyn alustaan, kuten laktoosia., Näitä entsyymejä tarvitaan vain silloin, kun kyseistä substraattia on saatavilla, joten operonien ilmentyminen indusoidaan tyypillisesti vain substraatin läsnä ollessa.
trp Operon: A Repressible Operon
E. coli voi syntetisoida tryptofaani käyttämällä entsyymejä, jotka ovat koodattu viisi rakenteelliset geenit sijaitsevat vierekkäin trp operon (Kuva 2). Kun ympäristön tryptofaani on alhainen, operoni käynnistetään. Tämä tarkoittaa, että transkriptio aloitetaan, geenit ilmaistaan ja tryptofaani syntetisoidaan., Jos tryptofaania kuitenkin esiintyy ympäristössä, trp-operoni sammutetaan. Transkriptiota ei tapahdu eikä tryptofaania syntetisoida.
Kun tryptofaania ei ole läsnä solu, repressori itse ei sitoudu operaattori; siksi operon on aktiivinen ja tryptofaani on syntetisoitu. Kuitenkin, kun tryptofaania kerääntyy soluun, kaksi tryptofaani molekyylit sitoutuvat trp repressori-molekyyli, joka muuttaa muotoaan, jolloin se voi sitoutua trp-operaattori., Tämä sitoutuminen aktiivinen muoto trp repressori operaattori estää RNA-polymeraasi alkaen litterointi rakenteelliset geenit, pysäyttää ilmaus operon. Siten operonin ohjaaman biosynteettisen reitin varsinainen tuote säätelee operonin ilmentymistä.

Kuva 2. Tryptofaanin syntetisoimiseen E. coli-bakteerissa tarvittavat viisi rakenteellista geeniä sijaitsevat vierekkäin trp operonissa. Kun tryptofaania ei ole, repressoriproteiini ei sitoudu toimijaan, ja geenit transkriptoidaan., Kun tryptofaania on runsaasti, tryptofaani sitoo repressori proteiinia operaattorilla järjestyksessä. Tämä estää fyysisesti RNA-polymeraasia tryptofaanin biosynteesigeenien transkriboimisesta.
Katso video saadaksesi lisätietoa trp operonista.
lac Operon: An Indusoituva Operon
lac-operonin on esimerkki indusoituva operon, että on myös edellyttää aktivointi ilman glukoosia (Kuva 3)., Lac operon koodaa kolme rakenteelliset geenit tarpeen hankkia ja käsitellä disakkaridi on laktoosi-ympäristöstä, murtaa se alas yksinkertaisia sokereita glukoosia ja galaktoosia. Jotta lac operon voidaan ilmaista, laktoosin on oltava läsnä. Tämä on solulle järkevää, koska olisi energeettisesti tuhlaavaa luoda entsyymit käsittelemään laktoosia, jos laktoosia ei olisi saatavilla.
puuttuessa laktoosi, lac repressori on sidottu operaattorin alueella lac operon, fyysisesti estää RNA-polymeraasi alkaen litterointi rakenteelliset geenit., Laktoosin läsnä ollessa solun sisällä oleva laktoosi kuitenkin muuntuu allolaktoosiksi. Allolactose toimii induktori molekyyli, sitoutuminen repressori ja muuttaa sen muodon, niin että se ei enää pysty sitoutumaan operaattori DNA. Poistaminen repressori läsnä laktoosin avulla RNA-polymeraasi liikkua toimija alueella ja aloittaa transkriptio lac rakenteelliset geenit.
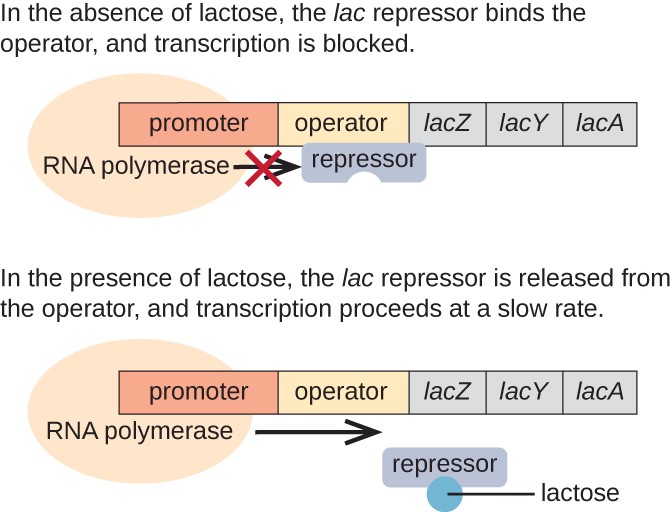
Kuva 3. Kolme rakenteellista geeniä, joita tarvitaan E. colin laktoosin hajottamiseen, sijaitsevat vierekkäin lac operonissa., Kun laktoosi on poissa, repressori proteiini sitoutuu operaattori, fyysisesti estää RNA-polymeraasi alkaen litterointi lac rakenteelliset geenit. Kun laktoosi on saatavilla, laktoosi-molekyyli sitoo repressori proteiinia, estää repressori sitoutumisen operaattori järjestyksessä, ja geenit ovat puhtaaksi.
lac Operon: Aktivointi Catabolite Activator Protein

Kuva 4. Kun E. coli kasvatetaan kahden substraatin läsnä ollessa, se käyttää suositeltua substraattia (tässä tapauksessa glukoosia), kunnes se on tyhjentynyt., Sitten toisen substraatin aineenvaihduntaan tarvittavat entsyymit ilmaistaan ja kasvu jatkuu, joskin hitaammin.
bakteereilla on tyypillisesti kyky käyttää erilaisia substraatteja hiililähteinä. Koska glukoosi on kuitenkin yleensä parempi kuin muut substraatit, bakteereilla on mekanismeja, joilla varmistetaan, että vaihtoehtoisia substraatteja käytetään vain silloin, kun glukoosi on loppunut. Lisäksi bakteereilla on mekanismeja, joilla varmistetaan, että entsyymejä vaihtoehtoisten substraattien käyttöön koodaavat geenit ilmaistaan vasta, kun vaihtoehtoinen substraatti on saatavilla., 1940-luvulla, Jacques Monod oli ensimmäinen osoittaa etusija tietyille alustoille yli muiden läpi hänen tutkimukset E. coli-bakteerin kasvua, kun viljellyt läsnä kaksi eri alustoille samanaikaisesti. Näissä tutkimuksissa syntyi diauksisia kasvukäyriä, kuten kuvassa 4. Vaikka ensisijainen substraatti glukoosia käytetään ensin, E. coli kasvaa nopeasti ja entsyymit laktoosi-aineenvaihduntaan puuttuvat. Kuitenkin, kun glukoosipitoisuus on tyhjentynyt, kasvu hidasta, asiakkuutta ilmaus entsyymejä tarvitaan aineenvaihduntaa toisen alustan, laktoosi., Huomaa, miten laktoosin kasvunopeus on hitaampi, kuten kasvukäyrän matalampi jyrkkyys osoittaa.
kyky vaihtaa glukoosi käyttää toiseen alustaan, kuten laktoosi-intoleranssi on seurausta toiminnan entsyymiä nimeltä Entsyymi II a (EIIA). Kun glukoosipitoisuus laskee, solut tuottavat vähemmän ATP: tä kataboliasta (KS.hiilihydraattien katabolia) ja EIIA fosforyloituu., Fosforyloitu EIIA aktivoi adenylyl cyclase, entsyymiä, joka muuntaa joitakin jäljellä ATP syklinen AMP (cAMP), syklinen johdannainen AMP ja tärkeä signalointi molekyyli mukana glukoosi-ja energia-aineenvaihduntaa E. coli. Tämän seurauksena leiritasot alkavat nousta solussa (kuva 5).

Kuva 5. Kun ATP-tasot laskevat, koska ehtyminen glukoosin, jotkut jäljellä ATP muunnetaan leirin adenylyl cyclase. Näin kohonneet leiritasot viestivät glukoosivajeesta.,
lac-operonin on myös tärkeä rooli tässä siirtyy käyttämään glukoosia käyttäen laktoosia. Kun glukoosia on niukasti, adenylyylisyklaasin aktiivisuuden lisääntymisen aiheuttama kertyvä leiri sitoutuu kataboliittiaktivaattoriproteiiniin (CAP), joka tunnetaan myös nimellä cAMP-reseptoriproteiini (CRP). Kompleksi sitoutuu lac operonin promoottorialueeseen (kuva 6). Sääntely-alueilla nämä operons, KORKKI sitoutumiskohta sijaitsee ylävirtaan RNA-polymeraasin sitoutumiskohta promoottorin., Cap-cAMP-kompleksin sitoutuminen tähän kohtaan Lisää RNA-polymeraasin sitoutumiskykyä promoottorin alueelle rakenteellisten geenien transkription aloittamiseksi. Näin ollen, siinä tapauksessa, että lac-operonin, sillä transkriptio tapahtuu, laktoosi on läsnä (poistamalla lac repressori proteiinia) ja glukoosipitoisuus on tyhjentynyt (jolloin sitova aktivoiva proteiini). Kun glukoosipitoisuus on korkea, on catabolite torjuntaan operons koodaus entsyymien aineenvaihdunnan vaihtoehto alustoille., Koska alhainen cAMP-tasoja näissä olosuhteissa, on riittämätön määrä CAP-leiri monimutkainen aktivoida transkriptio nämä operons. Taulukossa 1 on yhteenveto lac-operonin sääntelystä.

Kuva 6. (a) läsnäollessa leiri, KORKKI sitoo vetäjät operons, kuten lac operon, jotka koodaavat geenit entsyymien käyttöön vaihtoehtoinen alustoille. (b) lac-operonin ilmaistaan, on oltava aktivointi cAMP-CAP sekä poisto-lac repressori operaattorilta.,
| Taulukko 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
Miettiä
- Mikä vaikuttaa sitova trp operon repressori operaattori?
- miten ja milloin lac-repressoriproteiinin käyttäytyminen muuttuu?
- miten muuten lac operonia säännellään?
Global Vastauksia Prokaryooteissa
prokaryootit, on olemassa myös useita korkeampi geeni asetuksen, joka on kyky ohjata transkriptio monet liittyvät operons samanaikaisesti vastauksena ympäristön signaalin., Kaikkia samanaikaisesti ohjattuja operoneja kutsutaan reguloneiksi.
Alarmones
Kun sensing lähestyvästä stressiä, prokaryootit muuttaa ilmaus monenlaisia operons vastata koordinoidusti. He tekevät tämän tuottamalla alarmoneja, jotka ovat pieniä solunsisäisiä nukleotidijohdannaisia. Herätyskellot muuttavat sitä, mitkä geenit ilmentyvät ja stimuloivat tiettyjen stressireaktiogeenien ilmentymistä. Alarmonien käyttö geenien ilmentymisen muuttamiseen stressiin vastaamiseksi näyttää olevan tärkeää patogeenisille bakteereille., Kohdatessaan isännän puolustusmekanismit ja muut ankarat olosuhteet infektion aikana, monet virulenssigeenejä koodaavat operonit säätelevät herätyskellojen signalointia. Näiden vasteiden tuntemus on avain siihen, että voidaan täysin ymmärtää monien taudinaiheuttajien infektioprosessi ja kehittää hoitoja tämän prosessin torjumiseksi.,
Vaihtoehtoinen σ Tekijät
Koska σ alayksikköön bakteerien RNA-polymeraasi antaa spesifisyys siitä, mikä toteuttajien tulisi puhtaaksi, muuttamalla σ tekijä käyttää on toinen tapa, jolla bakteerit nopeasti ja globaalisti muuttaa mitä regulons ovat puhtaaksi tiettynä ajankohtana. Σ-tekijä tunnistaa sekvenssit bakteerin promoottorin sisällä, joten eri σ-tekijät tunnistavat kukin hieman erilaiset promoottorin sekvenssit., Tällä tavalla, kun solu aistii erityiset ympäristöolosuhteet, se voi vastata muuttamalla joka σ tekijä, se ilmaisee, halventava vanhaa ja tuottaa uutta puhtaaksi operons koodaus geenejä, joiden tuotteet ovat hyödyllisiä uusien ympäristön kannalta hyvässä kunnossa. Esimerkiksi päästää itiöitä bakteerien sukujen Bacillus ja Clostridium (joka sisältää monia taudinaiheuttajia), ryhmä σ tekijöitä ohjaa ilmaus monet geenit, joita tarvitaan itiöiden vastauksena itiöiden stimuloivia signaaleja.,
ajattele sitä
- mikä on nimi, joka annetaan joukolle operoneja, joita voidaan säännellä ryhmänä?
- minkälainen ärsyke laukaisisi eri σ-tekijän transkription?
Muita Menetelmiä Asetuksen Bakteerit: Vaimennus ja Riboswitches
Vaikka useimmat geenin ilmentyminen on säännelty tasolla transkriptio aloittamista, prokaryootit, siellä ovat myös mekanismeja, joilla voidaan valvoa sekä loppuun kirjoitus sekä käännös samanaikaisesti., Niiden löytymisen jälkeen näiden mekanismien on osoitettu ohjaavan monien prokaryoottisten operonien transkription ja käännöksen valmistumista. Koska nämä mekanismit linkki asetukseen, transkriptio ja käännös suoraan, ne ovat nimenomaan prokaryootit, koska nämä prosessit ovat fyysisesti erotettu eukaryooteissa.
Yksi tällainen sääntelyjärjestelmä on vaimennus, jolloin toissijainen varsi-silmukka-rakenteet muodostettu 5′ pää mRNA on puhtaaksi määrittää, jos transkriptio täydellinen synteesi tämän mRNA tapahtuu ja jos tämä mRNA käytetään käännöstä., Yli transkription sorron mekanismi jo keskustelleet, vaimennus kontrolloi myös ilmaus trp operon E. coli (Kuva 7). Trp operon sääntely-alue sisältää johtava sekvenssi kutsutaan trpL välillä operaattori ja ensimmäinen rakenteellisen geenin, joka on neljä ulottuu RNA, joka voi emäsparin toistensa kanssa eri yhdistelminä. Kun terminaattori stem-loop muodostuu, transkriptio päättyy vapauttaen RNA-polymeraasia mRNA: sta., Kuitenkin, kun antiterminator varsi-silmukka muotoja, tämä estää muodostumista terminator varsi-silmukka, jotta RNA-polymeraasi voi puhtaaksi rakenteelliset geenit.
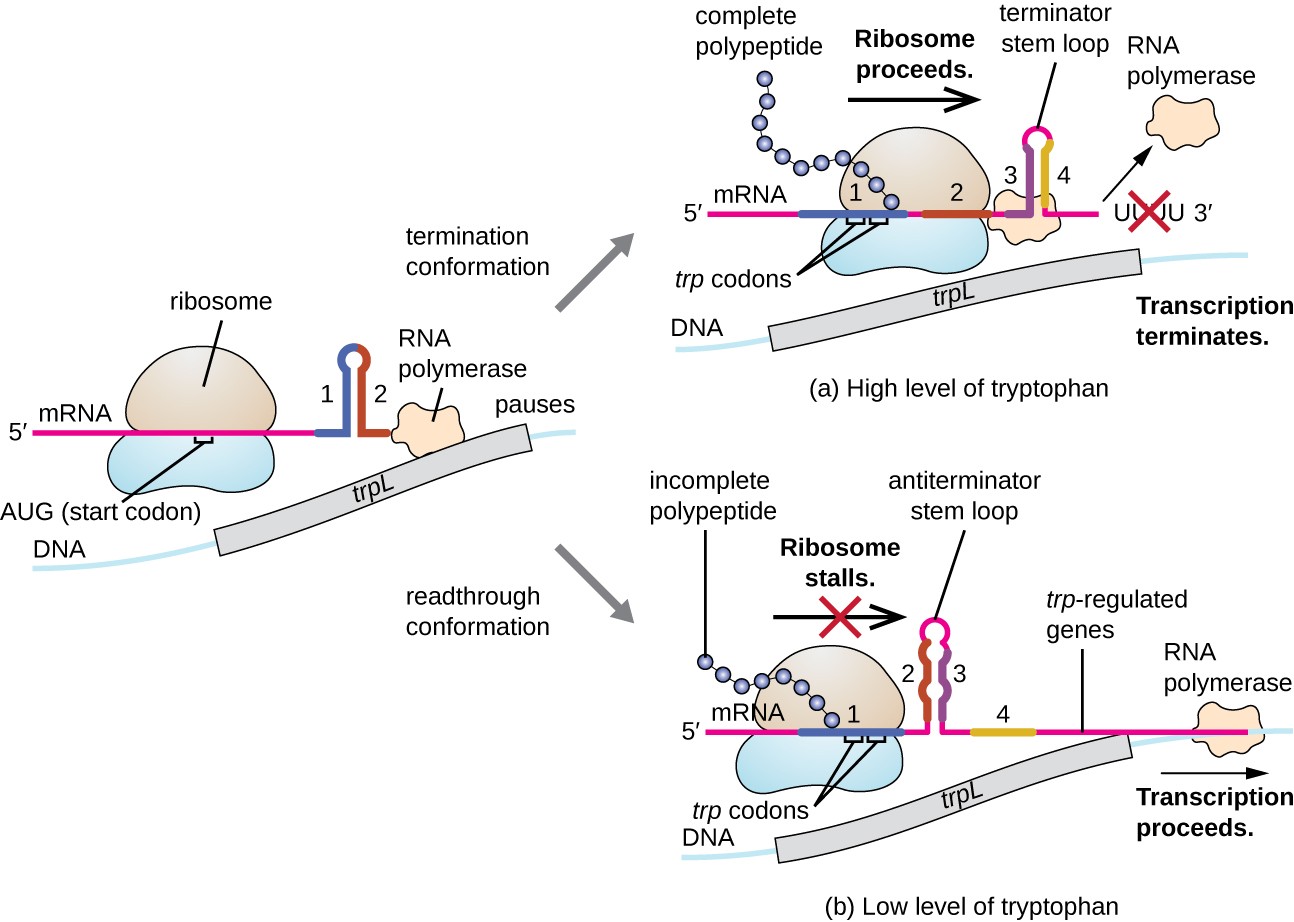
Kuva 7. Klikkaa nähdäksesi suuremman kuvan. Kun tryptofaania on runsaasti, käännös lyhyen johtaja peptidi koodattu trpL etenee, terminator-silmukka-alueiden välillä 3 ja 4 muotoja, ja transkriptio päättyy., Kun tryptofaani tasot ovat uhanalaisia, käännös lyhyen johtaja peptidi torit alue 1, jolloin alueiden 2 ja 3 muodostavat antiterminator silmukka, ja RNA-polymeraasi voi puhtaaksi rakenteelliset geenien trp operon.
liittyvä mekanismi samanaikaisesti asetuksen transkriptio ja käännös prokaryootit on käyttää riboswitch, pieni alue noncoding RNA löytyy sisällä 5′ lopussa noin prokaryootti-mRNA-molekyylejä (Kuva 8). Riboswitch saattaa sitoutua pieneen solunsisäiseen molekyyliin vakauttaakseen tietyt mRNA-molekyylin toissijaiset rakenteet., Pienen molekyylin sitoutuminen määrittää, mikä kantasilmukkarakenne muodostuu, mikä vaikuttaa mRNA-synteesin ja proteiinisynteesin loppuunsaattamiseen.

Kuva 8. Klikkaa isompaa kuvaa. Riboswitches löytynyt kuluessa prokaryootti-mRNA-molekyylit voivat sitoutua pieniä solunsisäisiä molekyylejä, vakauttaa tiettyjen RNA-rakenteita, jotka vaikuttavat joko loppuun synteesi mRNA-molekyyli itse (vasen) tai proteiini valmistettu käyttäen, että mRNA: n (oikealla).,
Muita Tekijöitä, jotka Vaikuttavat Geenien Ilmentymisen Prokaryooteissa ja Eukaryooteissa
Vaikka keskittyä keskusteluun transkription ohjaus käyttää prokaryoottiset operons esimerkkeinä, eukaryoottisesta transkription ohjaus on samanlainen monin tavoin. Kuten prokaryootit, eukaryoottisesta transkriptio voidaan ohjata sitova transkriptio tekijät, kuten repressors ja aktivaattoreita., Mielenkiintoista, eukaryoottisesta transkriptio voidaan vaikuttaa sitovia proteiineja alueille, DNA: n, nimeltään parantajia, melko kaukana geeni, DNA-silmukoiden helpottaa välillä tehostajana ja promoottori (Kuva 9). Kaiken kaikkiaan transkription sääntely on erittäin tehokas tapa kontrolloida geenien ilmentymistä sekä prokaryooteissa että eukaryooteissa. Kuitenkin, ohjaus geenien ilmentymisen eukaryooteissa vastauksena ympäristön ja solujen korostaa, voidaan tehdä muilla tavoin ilman sitovia transkriptio tekijät sääntelyn alueilla.,

Kuva 9. Eukaryooteissa tehostaja on transkriptiota edistävä DNA-sekvenssi. Jokainen tehostaja koostuu lyhyistä DNA-sekvensseistä, joita kutsutaan distaalisiksi ohjauselementeiksi. Aktivaattorit pakko distaalinen käyttöelementit vuorovaikutuksessa välittäjänä proteiineja ja transkriptio tekijät. Kahdella eri geenillä voi olla sama promoottori, mutta erilaiset distaaliset kontrollielementit, jotka mahdollistavat differentiaalisen geenin ilmentymisen.,
DNA-Tason säätö
eukaryooteissa DNA-molekyylejä tai siihen liittyvät histones voi olla kemiallisesti muutettu siten vaikuttaa transkription; tämä on nimeltään epigeneettiset asetuksella. Vaikka tiettyjen sytosiini nukleotidien DNA vastauksena ympäristötekijöiden on todettu vaikuttavan tällaisten DNA transkriptio, jossa DNA: n metylaatio yleisesti korreloivat alentaa tasoa geenien ilmentyminen., Lisäksi, vastauksena ympäristön tekijät, histoni-proteiinien pakkaus-DNA voi olla myös kemiallisesti muunnetut useilla eri tavoilla, mukaan lukien asetylointi ja deacetylation, vaikuttaa pakkaus-tila-DNA: ta ja siten vaikuttavat saatavuus löyhästi haavan DNA: n transkription. Nämä kemialliset muutokset voidaan joskus ylläpitää useita kierroksia solujen jakautumista, jolloin ainakin jotkut näistä epigeneettisiä muutoksia periytyvä.
tällä videolla kuvataan, miten epigeneettinen säätely säätelee geenien ilmentymistä.,
Miettiä
- Mikä estää tai mahdollistaa transkriptio edetä, kun vaimennus on käytössä?
- mikä määrittää riboswitchin tilan?
- kuvaa tehostimen toimintaa.
- kuvaa eukaryooteissa kahta epigeneettisen sääntelyn mekanismia.
Kliininen Painopiste: Travis, Resoluutio
Tässä esimerkissä todetaan, Travis on tarina, joka alkoi Toimintoja Geneettistä Materiaalia, RNA-Transkriptio, ja Miten Suvuton Prokaryootit Saavuttaa Geneettisen Monimuotoisuuden.,
Vaikka Travis selvisi hänen bout nekrotisoiva faskiitti, hän olisi nyt tehtävä ihon-varttaminen leikkaus, jonka jälkeen pitkän aikavälin fysioterapia. Perustuu määrä lihasmassaa hän menetti, on epätodennäköistä, että hänen jalkansa palaa täynnä voimaa, mutta hänen fyysinen terapeutin on optimistinen, että hän tulee takaisin joitakin käyttää hänen jalka.
laboratoriokokeet paljastivat, että Travisin infektion aiheuttaja oli A-ryhmän streptokokki (A-ryhmän streptokokki)., Lain edellyttämällä tavalla Travisin tapaus raportoitiin osavaltion terveysministeriölle ja lopulta tautien valvonta-ja Ehkäisykeskuksille (CDC). CDC: ssä Travisista eristetyn A-ryhmän kanta analysoitiin tarkemmin metisilliiniresistenssin varalta.
Metisilliiniresistenssi on geneettisesti koodattu ja yleistyy ryhmässä A strep horisontaalisen geeninsiirron kautta. Necrotizing fasciitis, veren virtaus tartunta-alue on yleensä rajoitettu, koska toiminnan eri geneettisesti koodattu bakteeri-myrkkyjä., Tämän vuoksi viiltotestin seurauksena verenvuotoa on tyypillisesti vähän tai ei lainkaan. Valitettavasti nämä bakteeri-myrkkyjä rajoittaa tehokkuutta suonensisäisiä antibiootteja clearing infektio, ihon ja taustalla kudos, mikä tarkoittaa, että antibioottiresistenssi ei yksin selitä tehottomuus Travis on hoito. Kuitenkin suonensisäisen antibioottihoidon oli perusteltua auttaa minimoimaan mahdollisen lopputuloksen sepsis, mikä on yleinen tulos nekrotisoiva faskiitti., Läpi genomin analyysi CDC: n kanta eristetty Travis, useita tärkeitä virulenssi geenejä olivat osoittautuneet koodattu prophages, mikä osoittaa, että transduktio on tärkeä horisontaalinen geenin siirtyminen näiden geenien yhdestä bakteeri solusta toiseen.
keskeiset käsitteet ja tiivistelmä
- geeniekspressio on tiukasti säännelty prosessi.
- geeniekspressio prokaryooteissa on pitkälti säädeltyä transkriptiopisteessä. Geeniekspressio eukaryooteissa on lisäksi säännelty jälkijättöisesti.,
- Prokaryoottiset rakenteelliset geenien liittyvä toiminta on usein organisoitu operons, kaikki määräysvallassa transkriptio yhdestä promoottori. Operonin sääntelyalueeseen kuuluvat järjestäjä itse ja promoottoria ympäröivä alue, johon transkriptiotekijät voivat sitoutua vaikuttamaan transkriptioon.
- Vaikka jotkut operons ovat konstitutiivisesti, useimmat sovelletaan asetuksen avulla transkriptio tekijät (repressors ja aktivaattorit)., A repressori sitoutuu operaattorin, DNA-sekvenssin sisällä sääntely-alueen välillä RNA-polymeraasin sitoutumiskohta promoottorin ja ensimmäisen rakenteellisen geenin, jolloin fyysisesti estää transkriptio nämä operons. Aktivaattori sitoutuu operonin säätelyalueella auttaen RNA-polymeraasia sitoutumaan promoottoriin, mikä tehostaa tämän operonin transkriptiota. Induktori vaikuttaa transkriptioon vuorovaikutuksessa repressorin tai aktivaattorin kanssa.
- trp operon on klassinen esimerkki repressible operon., Kun tryptofaani kertyy, tryptofaani sitoutuu repressori, joka sitten sitoutuu operaattori, estää edelleen transkriptio.
- lac operon on klassinen esimerkki indusoitavasta operonista. Kun solussa on laktoosia, se muuttuu allolaktoosiksi. Allolactose toimii induktori, sitoutuminen repressori ja repressori estää sitoutumisen operaattori. Tämä mahdollistaa rakenteellisten geenien transkription.
- lac operon on myös aktivoitavissa., Glukoosipitoisuuden ehtyessä osa solutason ATP: stä muuttuu campiksi, joka sitoutuu kataboliittiaktivaattoriproteiiniin (CAP). CAMP-CAP-kompleksi aktivoi Lac operonin transkription. Kun glukoosipitoisuus on korkea, sen läsnäolo estää transkriptio lac-operonin ja muut operons, jonka catabolite sortoa.
- Pieniä solunsisäisiä molekyylejä kutsutaan alarmones on tehty vastauksena eri ympäristön häiriötekijöille, jolloin bakteerit hallita transkriptio ryhmä operons, kutsutaan regulonin.,
- Bakteerit on kyky muuttaa joka σ tekijä RNA-polymeraasi he käyttävät vastauksena ympäristön olosuhteet nopeasti ja globaalisti muutos, joka regulons ovat puhtaaksi.
- Prokaryootit ovat sääntelyn mekanismeja, kuten vaimennus ja käyttö riboswitches, ohjata samanaikaisesti loppuun transkriptio ja käännös, että transkriptio. Nämä mekanismit toimivat muodostamalla runkosilmukoita parhaillaan syntetisoitavan mRNA-molekyylin 5 ’ päähän.,
- prokaryooteissa ja eukaryooteissa on muitakin geeniekspression säätelypisteitä. Eukaryooteissa epigeneettinen säätely DNA: n tai histonien kemiallisella muuntamisella ja RNA-käsittelyn säätely ovat kaksi menetelmää.
Useita Valinta
On operon geenien koodaavat entsyymejä biosynteettinen reitti on todennäköisesti mikä seuraavista?
- indusoituva
- repressible
- koostuu
- monocistronic
On operon koodaus geenejä, jotka ovat litteroitiin ja käännetty jatkuvasti tarjota solun jatkuva keskitason tasoilla proteiini tuotteita on sanottu olevan, mikä seuraavista?
- repressible
- indusoituva
- koostuu
- käytössä
Mikä seuraavista ehdoista johtaa maksimaalinen ilmentymä lac-operonin?
- laktoosi läsnä, glukoosi poissa
- laktoosi läsnä, glukoosin läsnä
- laktoosi-poissa -, glukoosi-poissa,
- laktoosi poissa, glukoosin läsnä
Mikä seuraavista on eräänlainen asetuksen geenien ilmentymisen ainutlaatuinen eukaryooteissa?,
- vaimennus
- käyttää vaihtoehtoista σ tekijä
- kemialliset muuttaminen histones
- alarmones
Täytä Tyhjä
DNA-sekvenssi, johon repressors voi sitoa, joka sijaitsee välillä promoottorin ja ensimmäisen rakenteellisia geeni on nimeltään ________.,
ehkäisyyn ilmaus operons koodaus alustan käyttää väyliä substraattien muut kuin glukoosia, kun glukoosi on läsnä, on nimeltään _______.
Miettiä
- Mitkä ovat kaksi tapaa, että bakteerit voivat vaikuttaa transkription useita eri operons samanaikaisesti vastauksena erityisesti ympäristön tilasta?
- seuraavassa kuvassa on Monod on alkuperäinen työ, joka on diauxic kasvu osoittaa kasvua E. coli yhtäaikainen läsnäolo ksyloosia ja glukoosi kuin vain hiilidioksidin lähteitä. Selittää, mitä on tapahtumassa pisteissä A–D-suhteessa hiilen lähteenä käytetään kasvuun, ja selittää, onko ksyloosi-käytä operon on ilmaissut (ja miksi)., Huomaa, että ksyloosin käyttöön tarvittavien entsyymien ilmentymistä säännellään samalla tavalla kuin laktoosin käyttöön tarvittavien entsyymien ilmentymistä.
