inlärningsmål
- jämför inducerbara operoner och repressibla operoner
- beskriv varför reglering av operoner är viktig
varje kärncell i en multicellulär organism innehåller kopior av samma DNA. På samma sätt innehåller alla celler i två rena bakteriekulturer inokulerade från samma startkoloni samma DNA, med undantag för förändringar som uppstår från spontana mutationer., Om varje cell i en multicellulär organism har samma DNA, hur är det då att celler i olika delar av organismens kropp uppvisar olika egenskaper? På samma sätt, hur är det att samma bakterieceller inom två rena kulturer som utsätts för olika miljöförhållanden kan uppvisa olika fenotyper? I båda fallen slår inte varje genetiskt identisk cell på, eller uttrycker, samma uppsättning gener. Endast en delmängd proteiner i en cell vid en given tidpunkt uttrycks.,
genomiskt DNA innehåller både strukturella gener, som kodar för produkter som fungerar som cellulära strukturer eller enzymer, och regulatoriska gener, som kodar för produkter som reglerar genuttryck. Uttrycket av en gen är en mycket reglerad process. Medan reglerande genuttryck i multicellulära organismer möjliggör cellulär differentiering, i encelliga organismer som prokaryoter, säkerställer det främst att en Cells resurser inte slösas bort vilket gör proteiner som cellen inte behöver vid den tiden.,
att belysa mekanismerna som styr genuttryck är viktigt för förståelsen av människors hälsa. Funktionsfel i denna process hos människor leder till utveckling av cancer och andra sjukdomar. Att förstå samspelet mellan ett patogens genuttryck och dess mänskliga värd är viktigt för förståelsen av en viss infektionssjukdom. Genreglering innebär ett komplext nät av interaktioner inom en given cell bland signaler från cellens miljö, signalmolekyler inom cellen och cellens DNA., Dessa interaktioner leder till uttryck av vissa gener och undertryckande av andra, beroende på omständigheterna.
prokaryoter och eukaryoter delar vissa likheter i deras mekanismer för att reglera genuttryck. men genuttryck i eukaryoter är mer komplicerat på grund av den tidsmässiga och rumsliga separationen mellan transkriptionsprocesserna och översättningen., Således, även om de flesta reglering av genuttryck sker genom transkriptionell kontroll i prokaryoter, reglering av genuttryck i eukaryoter sker på transkriptionsnivå och post-transkriptionellt (efter det primära transkriptet har gjorts).
prokaryotisk genreglering
i bakterier och archaea kodas strukturella proteiner med relaterade funktioner vanligtvis tillsammans i genomet i ett block som kallas en operon och transkriberas tillsammans under kontroll av en enda promotor, vilket resulterar i bildandet av ett polycistroniskt transkript (Figur 1)., På detta sätt kan reglering av transkriptionen av alla strukturella gener som kodar för de enzymer som katalyserar de många stegen i en enda biokemisk väg styras samtidigt, eftersom de antingen kommer att behövas samtidigt, eller ingen kommer att behövas. Till exempel, i E. coli, ligger alla strukturella gener som kodar enzymer som behövs för att använda laktos som en energikälla bredvid varandra i laktosoperonen (eller lac) under kontroll av en enda promotor, lac-promotorn., Franska forskare François Jacob (1920-2013) och Jacques Monod vid Pasteur-Institutet var den första att visa organisationen av bakteriella gener i operons, genom sina studier på lac operon av E. coli. För detta arbete vann de nobelpriset i fysiologi eller medicin 1965. Även om eukaryotiska gener inte är organiserade i operoner är prokaryotiska operoner utmärkta modeller för att lära sig om genreglering i allmänhet. Det finns vissa genkluster i eukaryoter som fungerar som operoner., Många av principerna kan tillämpas på eukaryota system och bidra till vår förståelse av förändringar i genuttryck i eukaryoter som kan leda till patologiska förändringar som cancer.

Figur 1. I prokaryoter organiseras strukturella gener av relaterad funktion ofta tillsammans på genomet och transkriberas tillsammans under kontroll av en enda promotor. Operatörens regulatoriska region omfattar både promotorn och operatören. Om en repressor binder till operatören, kommer de strukturella generna inte att transkriberas., Alternativt kan aktivatorer binda till regleringsområdet, vilket förbättrar transkriptionen.
varje operon innehåller DNA-sekvenser som påverkar dess egen transkription; dessa finns i en region som kallas regulatory region. Den regulatoriska regionen innefattar promotorn och regionen som omger promotorn, till vilken transkriptionsfaktorer, proteiner kodade av regulatoriska gener, kan binda. Transkriptionsfaktorer påverkar bindningen av RNA-polymeras till promotorn och tillåter dess progression att transkribera strukturella gener., En repressor är en transkriptionsfaktor som undertrycker transkription av en gen som svar på en yttre stimulans genom att binda till en DNA-sekvens inom den reglerande regionen som kallas operatören, som ligger mellan promotorns rna-polymerasbindningsställe och den transkriptionella startplatsen för den första strukturella genen. Repressor bindning blockerar fysiskt RNA-polymeras från transkribering av strukturella gener., Omvänt är en aktivator en transkriptionsfaktor som ökar transkriptionen av en gen som svar på en yttre stimulans genom att underlätta RNA-polymeras bindning till promotorn. En inducerare, en tredje typ av reglerande molekyl, är en liten molekyl som antingen aktiverar eller undertrycker transkription genom att interagera med en repressor eller en aktivator.
i prokaryoter finns det exempel på operoner vars genprodukter krävs ganska konsekvent och vars uttryck därför är oreglerat., Sådana operoner uttrycks konstitutionellt, vilket betyder att de transkriberas och översätts kontinuerligt för att ge cellen konstanta mellanliggande nivåer av proteinprodukterna. Sådana gener kodar enzymer som är involverade i hushållsfunktioner som krävs för cellulärt underhåll, inklusive DNA-replikering, reparation och uttryck, samt enzymer som är involverade i kärnmetabolism. Däremot finns det andra prokaryotiska operoner som uttrycks endast när det behövs och regleras av förtryckare, aktivatorer och inducerare.,
Tänk på det
- vad är delarna i DNA-sekvensen för en operon?
- vilka typer av reglerande molekyler finns det?
reglering genom Repression
prokaryota operoner kontrolleras vanligen genom bindning av repressorer till operatörsregioner, vilket förhindrar transkription av de strukturella generna. Sådana operoner klassificeras som antingen repressiva operoner eller inducerbara operoner. Repressibla operoner, som tryptofan (trp) operon, innehåller vanligtvis gener som kodar för enzymer som krävs för en biosyntetisk väg., Så länge produkten av vägen, som tryptofan, fortsätter att krävas av cellen, kommer en repressibel operon att fortsätta att uttryckas. Men när produkten av den biosyntetiska vägen börjar ackumuleras i cellen, tar bort behovet av att cellen fortsätter att göra mer, undertrycks uttrycket av operonen. Omvänt innehåller inducerbara operoner, som Lac-operonen av E. coli, ofta gener som kodar för enzymer i en väg som är involverad i metabolismen av ett specifikt substrat som laktos., Dessa enzymer krävs endast när det substratet är tillgängligt, vilket innebär att uttrycket av operonerna normalt induceras endast i närvaro av substratet.
trp Operon: En Repressible Operon
E. coli kan syntetisera tryptofan med hjälp av enzymer som kodas av fem strukturella gener som ligger bredvid varandra i trp operon (Figur 2). När miljö tryptofan är låg, är operonen påslagen. Detta innebär att transkription initieras, generna uttrycks och tryptofan syntetiseras., Om tryptofan är närvarande i miljön är trp-operonen avstängd. Transkription sker inte och tryptofan syntetiseras inte.
När tryptofan inte är närvarande i cellen binder repressorn i sig inte till operatören.därför är operonen aktiv och tryptofan syntetiseras. Men när tryptofan ackumuleras i cellen binder två tryptofanmolekyler till trp-repressormolekylen, som ändrar sin form, så att den kan binda till trp-operatören., Denna bindning av den aktiva formen av trp-repressorn till operatören blockerar RNA-polymeras från att transkribera de strukturella generna, stoppa uttrycket av operonen. Således reglerar den faktiska produkten av den biosyntetiska vägen som styrs av operonen uttrycket av operonen.

Figur 2. De fem strukturella generna som behövs för att syntetisera tryptofan i E. coli ligger bredvid varandra i trp-operonen. När tryptofan är frånvarande binder repressorproteinet inte till operatören, och generna transkriberas., När tryptofan är rikligt binder tryptofan repressorproteinet vid operatörssekvensen. Detta blockerar fysiskt RNA polymeras från transkribera tryptofan biosyntes gener.
titta på den här videon för att lära dig mer om trp-operonen.
lac Operon: en inducerbar Operon
lac operon är ett exempel på en inducerbar operon som också är föremål för aktivering i frånvaro av glukos (Figur 3)., Lac operon kodar för tre strukturella gener som är nödvändiga för att förvärva och bearbeta disackarid laktos från miljön, bryta ner den i enkla sockerarter glukos och galaktos. För att lacoperonen ska uttryckas måste laktos vara närvarande. Detta är meningsfullt för cellen eftersom det skulle vara energiskt slösaktigt att skapa enzymerna för att bearbeta laktos om laktos inte var tillgänglig.
i frånvaro av laktos är lac-repressorn bunden till operatörsområdet för lac-operonen, vilket fysiskt förhindrar RNA-polymeras från att transkribera de strukturella generna., Men när laktos är närvarande omvandlas laktos inuti cellen till allolaktos. Allolaktos tjänar som en inducermolekyl, bindning till repressorn och förändring av dess form så att den inte längre kan binda till operatorns DNA. Avlägsnande av repressorn i närvaro av laktos tillåter RNA-polymeras att röra sig genom operatörsområdet och börja transkribera av Lac strukturella gener.
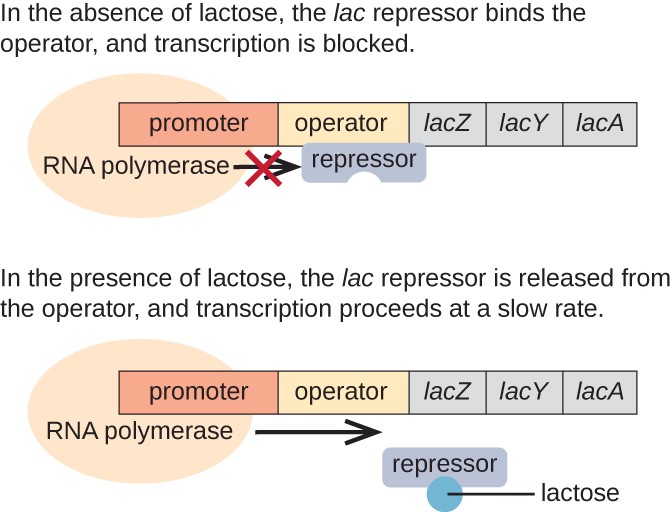
Figur 3. De tre strukturella generna som behövs för att försämra laktos i E. coli ligger bredvid varandra i lac-operonen., När laktos saknas binder repressorproteinet till operatören, vilket fysiskt blockerar RNA-polymerasen från att transkribera Lac strukturella gener. När laktos är tillgängligt binder en laktosmolekyl repressorproteinet, förhindrar repressorn från att binda till operatörssekvensen och generna transkriberas.
lac Operon: aktivering av Katabolit aktivator Protein

Figur 4. När den odlas i närvaro av två substrat använder E. coli det föredragna substratet (i detta fall glukos) tills det är utarmat., Därefter uttrycks enzymer som behövs för metabolismen av det andra substratet och tillväxten återupptas, men i långsammare takt.
bakterier har vanligtvis förmågan att använda en mängd olika substrat som kolkällor. Eftersom glukos vanligtvis är att föredra framför andra substrat har bakterier mekanismer för att säkerställa att alternativa substrat endast används när glukos har utarmat. Dessutom har bakterier mekanismer för att säkerställa att generna som kodar för enzymer för användning av alternativa substrat uttrycks endast när det alternativa substratet är tillgängligt., På 1940-talet var Jacques Monod den första som demonstrerade preferensen för vissa substrat jämfört med andra genom sina studier av E. colis tillväxt när den odlades i närvaro av två olika substrat samtidigt. Sådana studier genererade diauxiska tillväxtkurvor, som den som visas i Figur 4. Även om det föredragna substratet glukos används först växer E. coli snabbt och enzymerna för laktosmetabolism är frånvarande. Men när glukosnivåerna är utarmade, tillväxttakten långsam, inducera uttrycket av de enzymer som behövs för metabolismen av det andra substratet, laktos., Lägg märke till hur tillväxthastigheten i laktos är långsammare, vilket indikeras av tillväxtkurvans lägre branthet.
förmågan att byta från glukosanvändning till ett annat substrat som laktos är en följd av aktiviteten hos ett enzym som kallas enzymet IIA (EIIA). När glukosnivåerna sjunker producerar cellerna mindre ATP från katabolism (se katabolism av kolhydrater) och EIIA blir fosforylerad., Fosforylerad EIIA aktiverar adenylylcyklas, ett enzym som omvandlar några av de återstående ATP till cyklisk AMP (cAMP), ett cykliskt derivat av AMP och viktig signalmolekyl involverad i glukos och energimetabolism i E. coli. Som ett resultat börjar lägernivåerna stiga i cellen (Figur 5).

Figur 5. När ATP-nivåerna minskar på grund av utarmning av glukos omvandlas vissa återstående ATP till cAMP av adenylylylcyklas. Således signalerar ökade cAMP-nivåer glukosutarmning.,
Lac operon spelar också en roll i denna övergång från att använda glukos till att använda laktos. När glukos är knappa binder det ackumulerande lägret som orsakas av ökad adenylylyklasaktivitet till katabolitaktivatorprotein (CAP), även känt som cAMP-receptorprotein (CRP). Komplexet binder till promotorregionen i lac-operonen (Figur 6). I de regulatoriska regionerna av dessa operoner ligger en CAP-bindande plats uppströms rna-polymerasbindningsstället i promotorn., Bindning av CAP-cAMP-komplexet till denna webbplats ökar bindningsförmågan hos RNA-polymeras till promotorregionen för att initiera transkriptionen av de strukturella generna. I fallet med lac operon, för att transkription ska inträffa, måste laktos vara närvarande (avlägsnande av lac-repressorproteinet) och glukosnivåerna måste vara utarmade (vilket möjliggör bindning av ett aktiverande protein). När glukosnivåerna är höga finns katabolit förtryck av operoner som kodar för enzymer för metabolismen av alternativa substrat., På grund av låga lägernivåer under dessa förhållanden finns det inte tillräckligt med CAP-cAMP-komplexet för att aktivera transkription av dessa operoner. Se Tabell 1 för en sammanfattning av regleringen av lac-operonen.

Figur 6. (a) i närvaro av cAMP binder CAP till promotorer av operoner, som lac-operonen, som kodar för gener för enzymer för användning av alternativa substrat. b) för att lac-operonen ska kunna uttryckas måste aktiveringen ske med hjälp av cAMP-CAP samt avlägsnandet av lac-repressorn från operatören.,
| Tabell 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
Tänk på det
- vad påverkar bindningen av trp-operatörens repressor till operatören?
- hur och när förändras beteendet hos lac-repressorproteinet?
- förutom att vara repressibel, hur annars är Lac operon reglerad?
globala svar av prokaryoter
i prokaryoter finns det också flera högre nivåer av genreglering som har förmågan att styra transkriptionen av många relaterade operoner samtidigt som svar på en miljösignal., En grupp operoner som alla kontrolleras samtidigt kallas en regulon.
Alarmoner
vid avkänning av förestående stress förändrar prokaryoter uttrycket för en mängd olika operoner för att svara i samordning. De gör detta genom produktion av alarmoner, som är små intracellulära nukleotidderivat. Alarmoner förändrar vilka gener som uttrycks och stimulerar uttrycket av specifika stressresponsgener. Användningen av alarmoner för att förändra genuttryck som svar på stress verkar vara viktig för patogena bakterier., På möter värd försvarsmekanismer och andra svåra förhållanden under infektion, många operoner som kodar virulens gener uppregleras som svar på alarmon signalering. Kunskap om dessa svar är nyckeln till att fullt ut kunna förstå infektionsprocessen hos många patogener och till utvecklingen av terapier för att motverka denna process.,
alternativa σ-faktorer
eftersom σ-subenheten av bakteriellt RNA-polymeras ger specificitet om vilka promotorer som ska transkriberas, förändras σ-faktorn som används är ett annat sätt för bakterier att snabbt och globalt ändra vilka reguloner som transkriberas vid en given tidpunkt. Σ-faktorn känner igen sekvenser inom en bakteriell promotor, så olika σ-faktorer kommer var och en att känna igen lite olika promotorsekvenser., På så sätt, när cellen känner av specifika miljöförhållanden, kan den reagera genom att ändra vilken σ-faktor den uttrycker, försämra den gamla och producera en ny för att transkribera operonerna som kodar gener vars produkter kommer att vara användbara under det nya miljöförhållandet. Till exempel, i sporulerande bakterier av släktena Bacillus och Clostridium (som inkluderar många patogener), kontrollerar en grupp σ-faktorer uttrycket av de många gener som behövs för sporulering som svar på sporuleringsstimulerande signaler.,
Tänk på det
- vad är namnet på en samling operoner som kan regleras som en grupp?
- vilken typ av stimulans skulle utlösa transkriptionen av en annan σ-faktor?
ytterligare metoder för reglering av bakterier: dämpning och Riboswitches
även om de flesta genuttryck regleras i nivå med transkriptionsinitiering i prokaryoter, finns det också mekanismer för att kontrollera både slutförandet av transkription och översättning samtidigt., Sedan deras upptäckt har dessa mekanismer visat sig kontrollera slutförandet av transkription och översättning av många prokaryota operoner. Eftersom dessa mekanismer kopplar regleringen av transkription och översättning direkt, är de specifika för prokaryoter, eftersom dessa processer är fysiskt åtskilda i eukaryoter.
ett sådant regleringssystem är dämpning, varigenom sekundära stam-loop strukturer bildas inom 5 ’ slutet av en mRNA transkriberas avgöra om transkription för att slutföra syntesen av denna mRNA kommer att inträffa och om detta mRNA kommer att användas för översättning., Utöver den transkriptionella repressionsmekanismen som redan diskuterats kontrollerar dämpningen också uttrycket av trp-operonen i E. coli (Figur 7). Den trp operon rättsliga området innehåller en ledare sekvens som kallas trpL mellan operatören och den första strukturella genen, som har fyra sträckor av RNA som bas kan kopplas ihop med varandra i olika kombinationer. När en terminator stam-loop former, transkription avslutas, släppa RNA polymeras från mRNA., Men när en antiterminatorstamslinga bildas, förhindrar detta bildandet av terminatorstamslingan, så RNA-polymeras kan transkribera de strukturella generna.
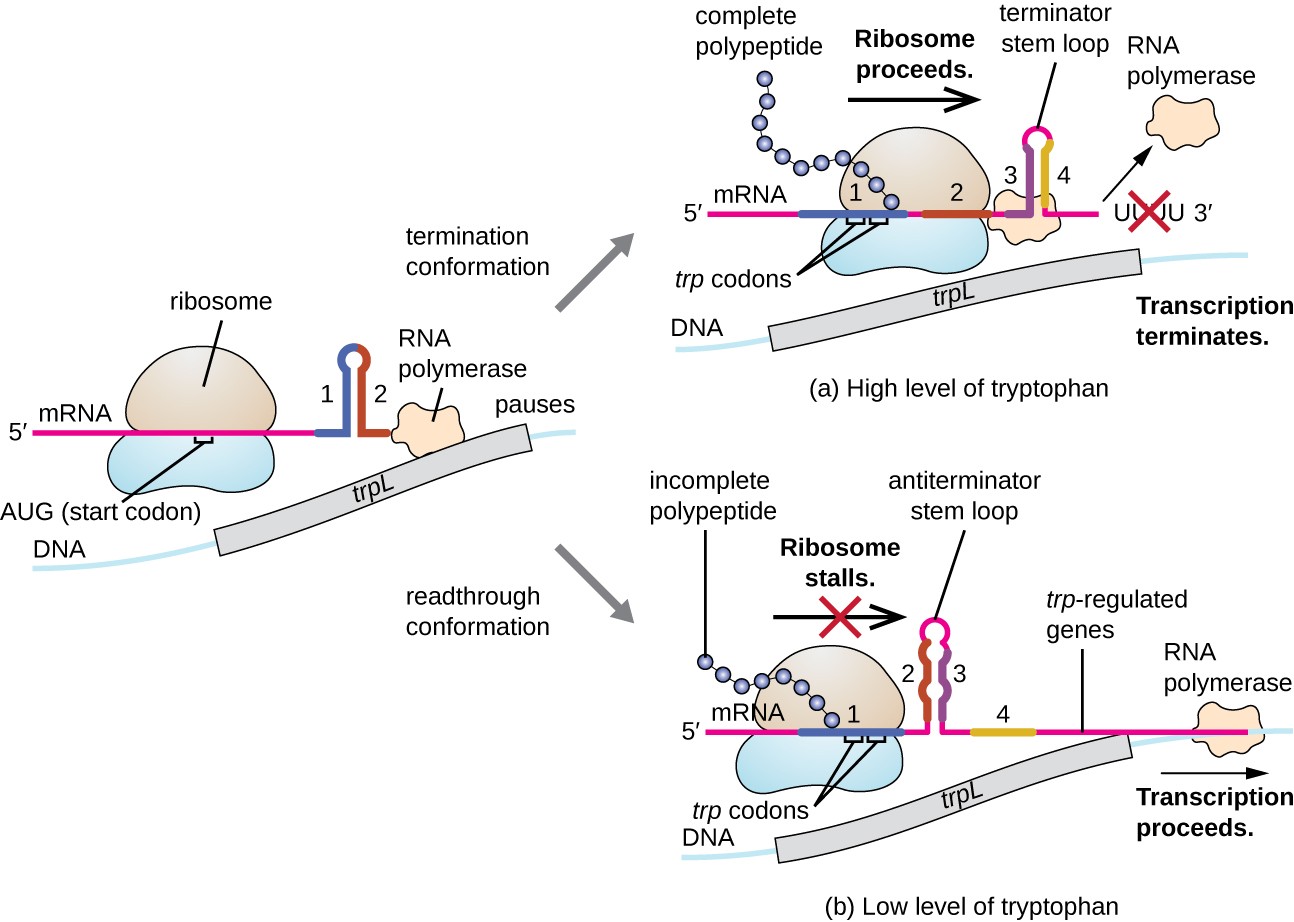
Figur 7. Klicka för att visa en större bild. När tryptofan är riklig, översättning av den korta leader peptiden kodad av trpL fortsätter terminatorslingan mellan regionerna 3 och 4 former och transkription avslutas., När tryptofannivåer är utarmade, översättning av den korta leader peptid bås i region 1, vilket gör att regioner 2 och 3 för att bilda en antiterminator slinga, och RNA polymeras kan transkribera de strukturella generna i trp operon.
en relaterad mekanism för samtidig reglering av transkription och översättning i prokaryoter är användningen av en riboswitch, en liten region av icke-kodande RNA som finns inom 5-änden av vissa prokaryotiska mRNA-molekyler (figur 8). En riboswitch kan binda till en liten intracellulär molekyl för att stabilisera vissa sekundära strukturer i mRNA-molekylen., Bindningen av den lilla molekylen bestämmer vilken stam-loop struktur som bildas, vilket påverkar slutförandet av mRNA-syntes och proteinsyntes.

figur 8. Klicka för en större bild. Riboswitches som finns inom prokaryota mRNA-molekyler kan binda till små intracellulära molekyler, stabilisera vissa RNA-strukturer, vilket påverkar antingen slutförandet av syntesen av mRNA-molekylen själv (vänster) eller proteinet som tillverkas med hjälp av den mRNA (höger).,
andra faktorer som påverkar genuttryck i prokaryoter och eukaryoter
även om fokus på vår diskussion om transkriptionskontroll används prokaryotiska operoner som exempel, är eukaryotisk transkriptionskontroll liknande på många sätt. Liksom i prokaryoter kan eukaryotisk transkription kontrolleras genom bindning av transkriptionsfaktorer, inklusive repressorer och aktivatorer., Intressant kan eukaryotisk transkription påverkas av bindningen av proteiner till DNA-regioner, kallade förstärkare, ganska långt ifrån genen, genom DNA-looping underlättad mellan förstärkaren och promotorn (Figur 9). Sammantaget är reglering av transkription ett mycket effektivt sätt att kontrollera genuttryck i både prokaryoter och eukaryoter. Kontrollen av genuttryck i eukaryoter som svar på miljö-och cellspänningar kan emellertid åstadkommas på ytterligare sätt utan bindning av transkriptionsfaktorer till regleringsregioner.,

Figur 9. I eukaryoter är en förstärkare en DNA-sekvens som främjar transkription. Varje förstärkare består av korta DNA-sekvenser som kallas distala kontrollelement. Aktivatorer som är bundna till de distala kontrollelementen interagerar med mediatorproteiner och transkriptionsfaktorer. Två olika gener kan ha samma promotor men olika distala kontrollelement, vilket möjliggör differentiell genuttryck.,
DNA-nivåkontroll
i eukaryoter kan DNA-molekylerna eller associerade histoner modifieras kemiskt på ett sådant sätt att de påverkar transkriptionen; detta kallas epigenetisk reglering. Metylering av vissa cytosinnukleotider i DNA som svar på miljöfaktorer har visat sig påverka användningen av sådant DNA för transkription, med DNA-metylering som vanligen korrelerar till sänkta nivåer av genuttryck., Dessutom, som svar på miljöfaktorer, histonproteiner för förpackning DNA kan också kemiskt modifieras på flera sätt, inklusive acetylering och deacetylering, påverka FÖRPACKNINGSTILLSTÅNDET av DNA och därmed påverka tillgången på löst sår DNA för transkription. Dessa kemiska modifieringar kan ibland upprätthållas genom flera rundor av celldelning, vilket gör åtminstone några av dessa epigenetiska förändringar ärftliga.
den här videon beskriver hur epigenetisk reglering styr genuttryck.,
Tänk på det
- vad stoppar eller tillåter transkription att fortsätta när dämpningen fungerar?
- vad bestämmer tillståndet för en riboswitch?
- beskriv funktionen hos en förstärkare.
- beskriv två mekanismer för epigenetisk reglering i eukaryoter.
kliniskt fokus: Travis, upplösning
detta exempel avslutar Travis berättelse som startade i funktionerna av genetiskt Material, rna transkription, och hur asexuella prokaryoter uppnå genetisk mångfald.,
Även om Travis överlevde sin bout med nekrotiserande fasciit, skulle han nu behöva genomgå en hudtransplantationskirurgi, följt av långvarig fysisk terapi. Baserat på den mängd muskelmassa han förlorade är det osannolikt att hans ben kommer tillbaka till full styrka, men hans fysioterapeut är optimistisk om att han kommer att återfå en viss användning av benet.
laboratorietestning avslöjade orsaksmedlet för Travis infektion var en stam av grupp A streptokocker (grupp A strep)., Enligt lag rapporterades Travis fall till statens hälsovårdsavdelning och slutligen till Centers for Disease Control and Prevention (CDC). Vid CDC analyserades stammen av grupp A strep isolerad från Travis mer noggrant för meticillinresistens.
meticillinresistens är genetiskt kodad och blir allt vanligare i Grupp A strep genom horisontell genöverföring. I nekrotiserande fasciit är blodflödet till det infekterade området typiskt begränsat på grund av verkan av olika genetiskt kodade bakterietoxiner., Det är därför det är typiskt liten eller ingen blödning som ett resultat av snittet testet. Tyvärr begränsar dessa bakterietoxiner effektiviteten av intravenösa antibiotika för att rensa infektion från huden och underliggande vävnad, vilket innebär att antibiotikaresistens ensam inte förklarar ineffektiviteten av Travis behandling. Ändå var intravenös antibiotikabehandling motiverad för att minimera det möjliga resultatet av sepsis, vilket är ett vanligt resultat av nekrotiserande fasciit., Genom genomisk analys av CDC av stammen isolerad från Travis visade sig flera av de viktiga virulensgenerna vara kodade på profetior, vilket indikerar att transduktion är viktig i den horisontella genöverföringen av dessa gener från en bakteriecell till en annan.
nyckelbegrepp och sammanfattning
- genuttryck är en tätt reglerad process.
- genuttryck i prokaryoter regleras i stor utsträckning vid transkriptionspunkten. Genuttryck i eukaryoter regleras dessutom post-transkriptionellt.,
- prokaryota strukturella gener av relaterad funktion är ofta organiserade i operoner, alla styrs av transkription från en enda promotor. Den regulatoriska regionen av en operon inkluderar promotorn själv och regionen som omger promotorn som transkriptionsfaktorer kan binda för att påverka transkription.
- även om vissa operoner är konstitutionellt uttryckta, är de flesta föremål för reglering genom användning av transkriptionsfaktorer (repressorer och aktivatorer)., En repressor binder till en operatör, en DNA-sekvens inom regleringsområdet mellan RNA-polymerasbindningsstället i promotorn och den första strukturella genen, vilket fysiskt blockerar transkriptionen av dessa operoner. En aktivator binder inom en operas regulatoriska region, vilket hjälper RNA-polymeras att binda till promotorn, vilket förbättrar transkriptionen av denna operon. En inducerare påverkar transkription genom att interagera med en repressor eller aktivator.
- trp operon är ett klassiskt exempel på en repressibel operon., När tryptofan ackumuleras binder tryptofan till en repressor, som sedan binder till operatören, vilket förhindrar ytterligare transkription.
- lac operon är ett klassiskt exempel en inducerbar operon. När laktos är närvarande i cellen omvandlas den till allolaktos. Allolaktos fungerar som en inducerare, bindande för repressorn och förhindrar repressorn från att binda till operatören. Detta möjliggör transkription av de strukturella generna.
- lac operon är också föremål för aktivering., När glukosnivåerna är utarmade omvandlas vissa cellulära ATP till cAMP, vilket binder till katabolitaktivatorproteinet (CAP). Lägret-CAP komplex aktiverar transkription av lac operon. När glukosnivåerna är höga förhindrar dess närvaro transkription av lac-operonen och andra operoner genom katabolit förtryck.
- små intracellulära molekyler som kallas alarmoner görs som svar på olika miljöbelastningar, vilket gör det möjligt för bakterier att kontrollera transkriptionen av en grupp operoner, kallad en regulon.,
- bakterier har förmågan att ändra vilken σ-faktor av RNA-polymeras de använder som svar på miljöförhållanden för att snabbt och globalt ändra vilka reguloner som transkriberas.
- prokaryoter har regleringsmekanismer, inklusive dämpning och användning av riboswitches, för att samtidigt kontrollera slutförandet av transkription och översättning från det transkriptet. Dessa mekanismer arbetar genom bildandet av stamslingor i 5-änden av en mRNA-molekyl som för närvarande syntetiseras.,
- Det finns ytterligare punkter för reglering av genuttryck i prokaryoter och eukaryoter. I eukaryoter är epigenetisk reglering genom kemisk modifiering av DNA eller histoner och reglering av RNA-bearbetning två metoder.
flervalsfrågor
en operon av gener som kodar för enzymer i en biosyntetisk väg kommer sannolikt att vara vilken av följande?
- inducerbar
- repressibel
- konstitutiv
- monocistronic
en operonkodningsgener som transkriberas och översätts kontinuerligt för att ge cellen konstanta mellanliggande nivåer av proteinprodukterna sägs vara vilken av följande?
- repressibel
- inducerbar
- konstitutiv
- aktiverad
vilka av följande villkor leder till maximalt uttryck för lac-operonen?
- laktos närvarande, glukos frånvarande
- laktos närvarande, glukos närvarande
- laktos frånvarande, glukos frånvarande
- laktos frånvarande, glukos närvarande
vilken av följande är en typ av reglering av genuttryck som är unik för eukaryoter?,
- dämpning
- användning av alternativa σ faktor
- kemisk modifiering av histoner
- alarmoner
fyll i blanketten
DNA-sekvensen, till vilken förtryckare kan binda, som ligger mellan promotorn och den första strukturella genen kallas_______.,
förebyggande av uttryck av operoner som kodar för substrat användning vägar för andra substrat än glukos när glukos är närvarande kallas______.
Tänk på det
- vad är två sätt att bakterier kan påverka transkriptionen av flera olika operoner samtidigt som svar på ett visst miljöförhållande?
- följande siffra är från Monods ursprungliga arbete med diauxisk tillväxt som visar tillväxten av E. coli i samtidig närvaro av xylos och glukos som de enda kolkällorna. Förklara vad som händer vid punkterna A-D med avseende på kolkällan som används för tillväxt och förklara om xylosanvändningsoperon uttrycks (och varför)., Observera att uttryck av de enzymer som krävs för användning av xylos regleras på ett sätt som liknar uttrycket av de enzymer som krävs för användning av laktos.
