læringsmål
- Sammenlign inducible operons og repressible operons
- Beskrive, hvorfor regulering af operons er vigtig
Hver kerneholdige celler i en flercellede organisme indeholder kopier af samme DNA. Tilsvarende indeholder alle celler i to rene bakteriekulturer inokuleret fra den samme startkoloni det samme DNA, med undtagelse af ændringer, der opstår som følge af spontane mutationer., Hvis hver celle i en multicellulær organisme har det samme DNA, hvordan er det så, at celler i forskellige dele af organismens krop udviser forskellige egenskaber? På samme måde, hvordan er det, at de samme bakterieceller inden for to rene kulturer udsat for forskellige miljøforhold kan udvise forskellige fænotyper? I begge tilfælde tænder eller udtrykker hver genetisk identisk celle ikke det samme sæt gener. Kun en delmængde af proteiner i en celle på et givet tidspunkt udtrykkes.,
genomisk DNA indeholder både strukturelle gener, som koder for produkter, der tjener som cellulære strukturer eller en .ymer, og regulatoriske gener, som koder for produkter, der regulerer genekspression. Ekspressionen af et gen er en stærkt reguleret proces. Mens regulering af genekspression i multicellulære organismer muliggør cellulær differentiering i enkeltcellede organismer som prokaryoter, sikrer det primært, at en celles ressourcer ikke spildes, hvilket gør proteiner, som cellen ikke har brug for på det tidspunkt.,
belysning af mekanismerne, der kontrollerer genekspression, er vigtig for forståelsen af menneskers sundhed. Fejl i denne proces hos mennesker fører til udvikling af kræft og andre sygdomme. Forståelse af interaktionen mellem genekspression af et patogen og dets menneskelige vært er vigtig for forståelsen af en bestemt infektionssygdom. Genregulering involverer en kompleks bane af interaktioner inden for en given celle blandt signaler fra cellens miljø, signalering molekyler i cellen, og cellens DNA., Disse interaktioner fører til ekspression af nogle gener og undertrykkelse af andre afhængigt af omstændighederne.prokaryoter og eukaryoter deler nogle ligheder i deres mekanismer til regulering af genekspression; imidlertid er genekspression i eukaryoter mere kompliceret på grund af den tidsmæssige og rumlige adskillelse mellem processerne for transkription og translation., Selvom de fleste regulering af genekspression forekommer gennem transkriptionel kontrol i prokaryoter, forekommer regulering af genekspression i eukaryoter på transkriptionsniveau og post-transkriptionelt (efter at det primære transkription er foretaget).
Prokaryote Gen Forordning
I bakterier og arkæer, strukturelle proteiner med relateret funktioner, der normalt kodet sammen inden for genomet i en blok, der kaldes en operon, og er transskriberet sammen under kontrol af en enkelt promotor, hvilket resulterer i dannelsen af en polycistronic udskrift (Figur 1)., På denne måde, regulering af transkription af alle de strukturelle gener, der koder for enzymer, der katalyserer de mange trin i en enkelt biokemisk pathway kan styres samtidigt, fordi de enten vil alle være behov for på samme tid, eller ingen vil være behov for. For eksempel i E. coli ligger alle de strukturelle gener, der koder for en .ymer, der er nødvendige for at bruge lactose som energikilde, ved siden af hinanden i lactosen (eller lac) operonen under kontrol af en enkelt promotor, lac-promotoren., Franske forskere François Jacob (1920-2013) og Jacques Monod ved Pasteur-Instituttet, var den første til at vise organisationen af bakterielle gener i operons, gennem deres studier på lac-operonen af E. coli. Til dette arbejde vandt de Nobelprisen i fysiologi eller medicin i 1965. Selvom eukaryote gener ikke er organiseret i operoner, er prokaryote operoner fremragende modeller til at lære om genregulering generelt. Der er nogle genklynger i eukaryoter, der fungerer som operoner., Mange af principperne kan anvendes på eukaryote systemer og bidrage til vores forståelse af ændringer i genekspression i eukaryoter, der kan resultere i patologiske ændringer som kræft.

Figur 1. I prokaryoter organiseres strukturelle gener af relateret funktion ofte sammen på genomet og transkriberes sammen under kontrol af en enkelt promotor. Operons reguleringsområde omfatter både promotoren og operatøren. Hvis en repressor binder til operatøren, vil de strukturelle gener ikke blive transkriberet., Alternativt kan aktivatorer binde til den regulerende region, øge transkription.
hver operon indeholder DNA-sekvenser, der påvirker sin egen transkription; disse er placeret i en region kaldet regulatory region. Den regulerende region omfatter promotoren og regionen omkring promotoren, som transkriptionsfaktorer, proteiner kodet af regulatoriske gener, kan binde. Transkriptionsfaktorer påvirker bindingen af RNA-polymerase til promotoren og tillader dens progression at transkribere strukturelle gener., En repressor er en transkriptionsfaktor, der undertrykker transkription af et gen som reaktion på en ekstern stimulus ved at binde til en DNA-sekvens inden for det regulatoriske område kaldet operatøren, som er placeret mellem RNA-polymerasebindingsstedet for promotoren og det transkriptionelle startsted for det første strukturelle gen. Repressorbinding blokerer fysisk RNA-polymerase fra transkribering af strukturelle gener., Omvendt er en aktivator en transkriptionsfaktor, der øger transkriptionen af et gen som respons på en ekstern stimulus ved at lette RNA-polymerasebinding til promotoren. En inducer, en tredje type regulatorisk molekyle, er et lille molekyle, der enten aktiverer eller undertrykker transkription ved at interagere med en repressor eller en aktivator.i prokaryoter er der eksempler på operoner, hvis genprodukter kræves ret konsekvent, og hvis udtryk derfor er ureguleret., Sådanne operoner udtrykkes konstitutivt, hvilket betyder, at de transkriberes og oversættes kontinuerligt for at tilvejebringe cellen med konstante mellemniveauer af proteinprodukterne. Sådanne gener koder for en .ymer involveret i husholdnings funktioner, der kræves til cellulær vedligeholdelse, herunder DNA-replikation, reparation og ekspression samt en .ymer involveret i kernemetabolisme. I modsætning hertil er der andre prokaryote operoner, der kun udtrykkes, når det er nødvendigt og reguleres af undertrykkere, aktivatorer og induktorer.,
tænk over det
- hvad er delene i DNA-sekvensen af en operon?
- hvilke typer regulatoriske molekyler er der?
regulering ved undertrykkelse
prokaryote operoner kontrolleres almindeligvis ved binding af repressorer til operatørregioner og forhindrer derved transkription af de strukturelle gener. Sådanne operoner klassificeres som enten undertrykkelige operoner eller inducerbare operoner. Repressible operoner, som tryptophan (trp) operon, indeholder typisk gener, der koder for en .ymer, der kræves til en biosyntetisk vej., Så længe produktet af vejen, som tryptophan, fortsat kræves af cellen, vil en undertrykkende operon fortsat blive udtrykt. Men når produktet af den biosyntetiske vej begynder at akkumulere i cellen, fjerner behovet for, at cellen fortsætter med at gøre mere, undertrykkes operonens udtryk. Omvendt indeholder inducerbare operoner, ligesom lac operon af E. coli, ofte gener, der koder for en .ymer i en vej involveret i metabolismen af et specifikt substrat som lactose., Disse en .ymer er kun nødvendige, når dette substrat er tilgængeligt, således induceres ekspression af operonerne typisk kun i nærvær af substratet.
trp Operon: en undertrykkende Operon
E. coli kan syntetisere tryptophan ved hjælp af en .ymer, der er kodet af fem strukturelle gener placeret ved siden af hinanden i trp operon (figur 2). Når miljøtryptophan er lav, er operonen tændt. Dette betyder, at transkription initieres, generne udtrykkes, og tryptophan syntetiseres., Men hvis tryptophan er til stede i miljøet, er trp operon slukket. Transskription forekommer ikke, og tryptophan syntetiseres ikke.
når tryptophan ikke er til stede i cellen, binder undertrykkeren i sig selv ikke til operatøren; derfor er operonen aktiv, og tryptophan syntetiseres. Når tryptophan imidlertid akkumuleres i cellen, binder to tryptophanmolekyler til trp-undertrykkelsesmolekylet, som ændrer dets form, så det kan binde til trp-operatøren., Denne binding af den aktive form af trp-undertrykkeren til operatøren blokerer RNA-polymerase fra at transkribere de strukturelle gener, hvilket stopper ekspressionen af operonen. Således regulerer det faktiske produkt af den biosyntetiske vej, der styres af operonen, ekspressionen af operonen.

Figur 2. De fem strukturelle gener, der er nødvendige for at syntetisere tryptophan i E. coli, er placeret ved siden af hinanden i trp-operonen. Når tryptophan er fraværende, binder represorproteinet ikke til operatøren, og generne transkriberes., Når tryptophan er rigeligt, binder tryptophan represorproteinet ved operatørsekvensen. Dette blokerer fysisk RNA-polymerasen fra at transkribere tryptophanbiosyntesegener.
se denne video for at lære mere om trp operon.
lac Operon: en inducerbar Operon
lac operon er et eksempel på en inducerbar operon, der også er genstand for aktivering i fravær af glucose (figur 3)., Lac operon koder for tre strukturelle gener, der er nødvendige for at erhverve og behandle disaccharid lactose fra miljøet, bryde det ned i den enkle sukker glucose og galactose. For at lac-operonen skal udtrykkes, skal lactose være til stede. Dette giver mening for cellen, fordi det ville være energisk spildt at skabe en .ymerne til at behandle lactose, hvis laktose ikke var tilgængelig.
i fravær af lactose er lac-undertrykkeren bundet til operatørområdet for lac-operonen, hvilket fysisk forhindrer RNA-polymerase i at transkribere de strukturelle gener., Når lactose er til stede, omdannes lactosen inde i cellen til allolactose. Allolactose tjener som et inducermolekyle, der binder til undertrykkeren og ændrer dens form, så den ikke længere er i stand til at binde til operatørens DNA. Fjernelse af undertrykkeren i nærvær af lactose tillader RNA-polymerase at bevæge sig gennem operatørområdet og begynde transkription af lac-strukturelle gener.
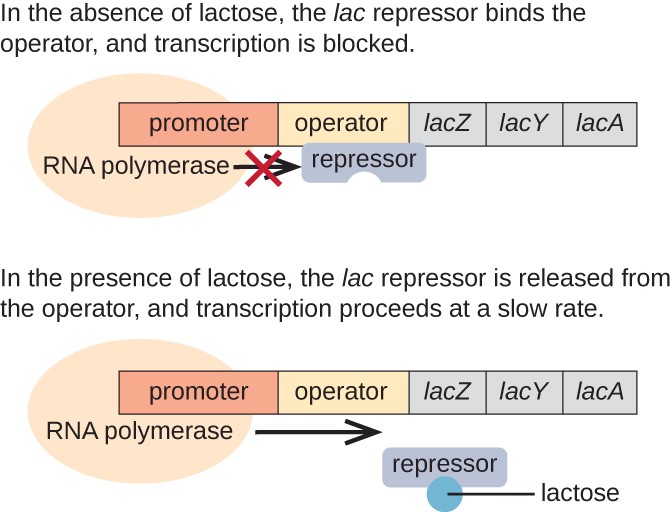
Figur 3. De tre strukturelle gener, der er nødvendige for at nedbryde lactose i E. coli, er placeret ved siden af hinanden i lac operon., Når lactose er fraværende, binder represorproteinet sig til operatøren, hvilket fysisk blokerer RNA-polymerasen fra at transkribere lac-strukturelle gener. Når lactose er tilgængelig, binder et lactosemolekyle represorproteinet, forhindrer represoren i at binde til operatørsekvensen, og generne transkriberes.
lac-Operonen: Aktivering af Catabolite Aktivator Protein

Figur 4. Når der dyrkes i nærvær af to substrater, bruger E. coli det foretrukne substrat (i dette tilfælde glucose), indtil det er udtømt., Derefter udtrykkes en .ymer, der er nødvendige for metabolismen af det andet substrat, og væksten genoptages, skønt i en langsommere hastighed.
bakterier har typisk evnen til at bruge en række substrater som carbonkilder. Da glukose normalt foretrækkes frem for andre substrater, har bakterier mekanismer til at sikre, at alternative substrater kun anvendes, når glukose er udtømt. Derudover har bakterier mekanismer til at sikre, at generne, der koder for en .ymer til anvendelse af alternative substrater, kun udtrykkes, når det alternative substrat er tilgængeligt., I 1940’erne, Jacques Monod var den første til at demonstrere til fordel for bestemte substrater over andre gennem sine studier af E. coli ‘ s vækst, når de dyrkes i nærværelse af to forskellige substrater samtidigt. Sådanne undersøgelser genererede diau .iske vækstkurver, som den, der er vist i figur 4. Selvom den foretrukne substratglukose først anvendes, vokser E. coli hurtigt, og en .ymerne til lactosemetabolisme er fraværende. Når først glukoseniveauerne er udtømt, er vækstraterne langsomme, hvilket fremkalder ekspressionen af de en .ymer, der er nødvendige for metabolismen af det andet substrat, lactose., Bemærk, hvordan vækstraten i lactose er langsommere, som indikeret af den lavere stejlhed i vækstkurven.
evnen til at skifte fra glukoseanvendelse til et andet substrat som lactose er en konsekvens af aktiviteten af et en .ym kaldet en .ym IIA (EIIA). Når glukoseniveauerne falder, producerer celler mindre ATP fra katabolisme (se katabolisme af kulhydrater), og EIIA bliver phosphoryleret., Fosforylerede EIIA aktiverer adenylyl cyclase, et enzym, der omdanner nogle af de resterende ATP til cyklisk AMP (cAMP), en cyklisk derivat af AMP og vigtige signaler molekyle, der er involveret i glukose og energi stofskifte i E. coli. Som et resultat begynder lejrniveauerne at stige i cellen (figur 5).

Figur 5. Når ATP-niveauerne falder på grund af udtømning af glukose, omdannes nogle resterende ATP til cAMP ved adenylylcyklase. Således signalerer øgede cAMP-niveauer glukoseudtømning.,
lac operon spiller også en rolle i denne skift fra at bruge glukose til at bruge lactose. Når glukose er knap, binder den akkumulerende cAMP forårsaget af øget adenylylcyklaseaktivitet til katabolitaktivatorprotein (CAP), også kendt som cAMP-receptorprotein (CRP). Komplekset binder til promotorområdet af lac operon (figur 6). I de regulatoriske regioner af disse operoner er et CAP-bindingssted placeret opstrøms for RNA-polymerasebindingsstedet i promotoren., Binding af CAP-cAMP-komplekset til dette sted øger BINDINGSEVNEN af RNA-polymerase til promotorområdet for at initiere transkriptionen af de strukturelle gener. I tilfælde af lac operon, for at transkription skal forekomme, skal lactose være til stede (fjernelse af lac-undertrykkelsesproteinet), og glukoseniveauerne skal udtømmes (tillader binding af et aktiverende protein). Når glukoseniveauerne er høje, er der katabolitundertrykkelse af operoner, der koder for en .ymer til metabolisme af alternative substrater., På grund af lave lejrniveauer under disse forhold er der en utilstrækkelig mængde af CAP-cAMP-komplekset til at aktivere transkription af disse operoner. Se tabel 1 for et resum.af reguleringen af lac operon.

Figur 6. (a) i nærværelse af cAMP binder CAP sig til promotorer af operoner, som lac-operonen, der koder gener for en .ymer til brug af alternative substrater. b) for at lac-operonen kan udtrykkes, skal der ske aktivering ved hjælp af cAMP-CAP samt fjernelse af lac-repressoren fra operatøren.,
| Tabel 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
tænk over det
- hvad påvirker bindingen af trp operon repressor til operatøren?
- hvordan og hvornår ændres adfærden af lac repressorproteinet?
- ud over at være undertrykkende, hvordan ellers er lac operon reguleret?
globale responser af prokaryoter
i prokaryoter er der også flere højere niveauer af genregulering, der har evnen til at kontrollere transkriptionen af mange beslægtede operoner samtidigt som reaktion på et miljøsignal., En gruppe operoner, der alle styres samtidigt, kaldes en regulon.
Alarmones
når man mærker forestående stress, ændrer prokaryoter udtrykket af en lang række operoner for at reagere i koordination. De gør dette gennem produktion af alarmones, som er små intracellulære nukleotid derivater. Alarmoner ændrer hvilke gener der udtrykkes og stimulerer ekspressionen af specifikke stress-Responsgener. Brugen af alarmones til at ændre genekspression som reaktion på stress synes at være vigtig i patogene bakterier., Når man støder på værtsforsvarsmekanismer og andre barske forhold under infektion, mange operoner, der koder for virulensgener, reguleres som reaktion på alarmone-signalering. Kendskab til disse svar er nøglen til at være i stand til fuldt ud at forstå infektionsprocessen hos mange patogener og til udviklingen af terapier for at imødegå denne proces.,
Alternative Factors-faktorer
da bacterial-underenheden af bakteriel RNA-polymerase giver specificitet med hensyn til, hvilke promotorer der skal transkriberes, er ændring af den anvendte factor-faktor en anden måde for bakterier hurtigt og globalt at ændre, hvilke reguloner der transkriberes på et givet tidspunkt. Σ-faktoren genkender sekvenser inden for en bakteriel promotor, så forskellige factors-faktorer genkender hver lidt forskellige promotorsekvenser., På denne måde, når cellen registrerer specifikke miljøforhold, kan den reagere ved at ændre hvilken factor-faktor den udtrykker, nedbryde den gamle og producere en ny til at transkribere operonerne, der koder for gener, hvis produkter vil være nyttige under den nye miljøtilstand. For eksempel kontrollerer en gruppe σ-faktorer i sporulerende bakterier af slægterne Bacillus og Clostridium (som inkluderer mange patogener) ekspressionen af de mange gener, der er nødvendige til sporulation som reaktion på sporulationsstimulerende signaler.,
tænk over det
- hvad er navnet givet til en samling operoner, der kan reguleres som en gruppe?
- hvilken type stimulus ville udløse transkriptionen af en anden factor-faktor?
Yderligere Metoder til Regulering i Bakterier: Dæmpning og Riboswitches
Selv om de fleste gen-ekspression reguleres af transkription indledningen i prokaryotes, der er også mekanismer til at styre både afslutningen af transskription samt oversættelse samtidigt., Siden deres opdagelse har disse mekanismer vist sig at kontrollere færdiggørelsen af transkription og oversættelse af mange prokaryote operoner. Fordi disse mekanismer forbinder reguleringen af transkription og oversættelse direkte, er de specifikke for prokaryoter, fordi disse processer er fysisk adskilt i eukaryoter.
Et sådant reguleringssystem dæmpning, hvorved sekundære stem-loop strukturer, der dannes i 5′ – enden af mRNA bliver transskriberet afgøre, om transskription for at fuldføre syntesen af dette mRNA vil forekomme, og hvis dette mRNA vil blive brugt til oversættelse., Ud over den transkriptionelle undertrykkelsesmekanisme, der allerede er diskuteret, kontrollerer dæmpning også ekspression af trp-operonen i E. coli (Figur 7). TRP operon regulatory region indeholder en ledersekvens kaldet trpL mellem operatøren og det første strukturelle gen, som har fire strækninger af RNA, der kan basepar med hinanden i forskellige kombinationer. Når en terminator stamsløjfe dannes, transkription ophører, frigiver RNA polymerase fra mRNA., Når en antiterminator-stamsløjfe dannes, forhindrer dette imidlertid dannelsen af terminator-stamsløjfen, så RNA-polymerase kan transkribere de strukturelle gener.
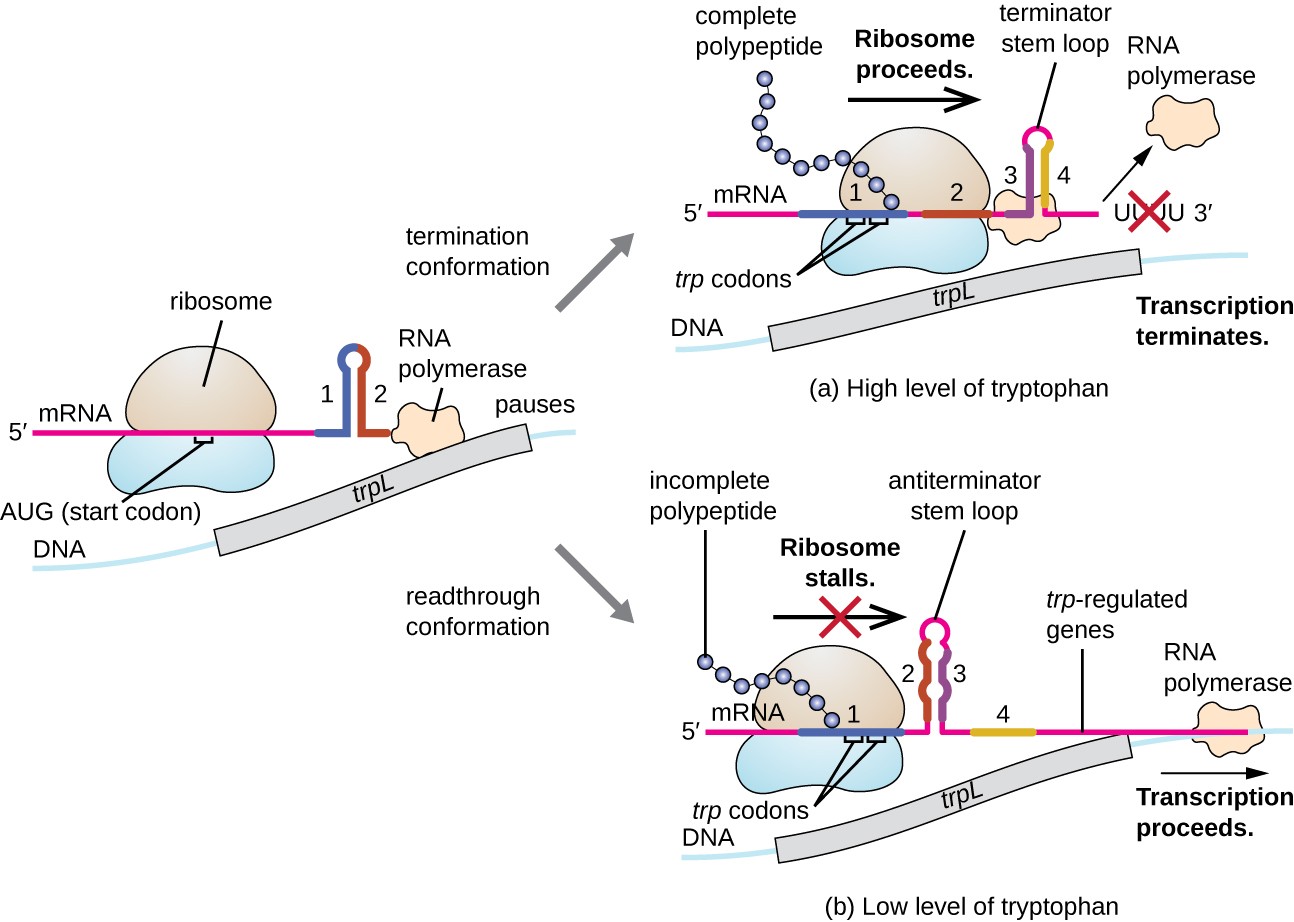
Figur 7. Klik for at se et større billede. Når tryptophan er rigeligt, oversættelse af den korte leder peptid kodet af trpL udbytte, terminator loop mellem regioner, 3 og 4 former, og transskription ophører., Når tryptofan niveauer er udtømte, oversættelse af den korte leder peptid boder på region 1, så regionerne får mulighed 2 og 3 for at danne en antiterminator loop, og RNA-polymerase kan transskribere de strukturelle gener af trp-operonen.
en relateret mekanisme for samtidig regulering af transkription og oversættelse i prokaryoter er brugen af en ribositchitch, et lille område af ikke-kodende RNA, der findes inden for 5′ – enden af nogle prokaryote mRNA-molekyler (figur 8). En ribositchitch kan binde til et lille intracellulært molekyle for at stabilisere visse sekundære strukturer af mRNA-molekylet., Bindingen af det lille molekyle bestemmer, hvilken stamsløjfestruktur der dannes, hvilket påvirker færdiggørelsen af mRNA-syntese og proteinsyntese.

Figur 8. Klik for større billede. Ribosititches, der findes inden for prokaryote mRNA-molekyler, kan binde til små intracellulære molekyler, stabilisere visse RNA-strukturer, hvilket påvirker enten færdiggørelsen af syntesen af selve mRNA-molekylet (venstre) eller proteinet fremstillet ved hjælp af det mRNA (højre).,
Andre Faktorer, der Påvirker genekspression i Prokaryotes og Eukaryoter
Selv om fokus på vores diskussion af transkriptionel kontrol anvendes prokaryote operons som eksempler, eukaryote transkriptionel kontrol ligner på mange måder. Som i prokaryoter kan eukaryotisk transkription styres gennem binding af transkriptionsfaktorer, herunder repressorer og aktivatorer., Interessant nok kan eukaryotisk transkription påvirkes af bindingen af proteiner til DNA-regioner, kaldet forstærkere, temmelig langt væk fra genet, gennem DNA-sløjfer, der letter mellem forstærkeren og promotoren (figur 9). Generelt er regulering af transkription en meget effektiv måde at kontrollere genekspression i både prokaryoter og eukaryoter. Imidlertid kan kontrollen med genekspression i eukaryoter som reaktion på miljømæssige og cellulære belastninger udføres på yderligere måder uden binding af transkriptionsfaktorer til regulerende regioner.,

Figur 9. I eukaryoter er en forstærker en DNA-sekvens, der fremmer transkription. Hver forstærker består af korte DNA-sekvenser kaldet distale kontrolelementer. Aktivatorer bundet til de distale kontrolelementer interagerer med mediatorproteiner og transkriptionsfaktorer. To forskellige gener kan have den samme promotor, men forskellige distale kontrolelementer, hvilket muliggør differentiel genekspression.,
DNA-niveau kontrol
i eukaryoter kan DNA-molekylerne eller tilknyttede histoner kemisk modificeres på en sådan måde, at de påvirker transkription; dette kaldes epigenetisk regulering. Methylering af visse cytosinnukleotider i DNA som reaktion på miljøfaktorer har vist sig at påvirke brugen af sådant DNA til transkription, med DNA-methylering, der ofte korrelerer med nedsatte niveauer af genekspression., Som reaktion på miljøfaktorer kan histonproteiner til emballering af DNA også ændres kemisk på flere måder, herunder acetylering og deacetylering, der påvirker emballagetilstanden for DNA og således påvirker tilgængeligheden af løst sår DNA til transkription. Disse kemiske modifikationer kan undertiden opretholdes gennem flere runder af celledeling, hvilket gør i det mindste nogle af disse epigenetiske ændringer arvelige.denne video beskriver, hvordan epigenetisk regulering styrer genekspression.,
tænk over det
- hvad stopper eller tillader transskription at fortsætte, når dæmpningen fungerer?
- hvad bestemmer tilstanden af en ribositchitch?
- beskriv funktionen af en forstærker.
- beskriv to mekanismer for epigenetisk regulering i eukaryoter.
klinisk fokus: Travis, opløsning
dette eksempel konkluderer Travis historie, der startede i funktionerne af genetisk materiale, RNA-transkription, og hvordan aseksuelle prokaryoter opnår genetisk mangfoldighed.,selvom Travis overlevede sin kamp med nekrotiserende fasciitis, skulle han nu gennemgå en hudtransplantationskirurgi efterfulgt af langvarig fysioterapi. Baseret på mængden af muskelmasse han tabte, er det usandsynligt, at hans ben vil vende tilbage til fuld styrke, men hans fysioterapeut er optimistisk, at han vil genvinde en vis brug af hans ben.
laboratorieundersøgelser afslørede, at det forårsagende middel til Travis ‘ infektion var en stamme af gruppe A streptococcus (gruppe A strep)., Som krævet i loven blev Travis sag rapporteret til statens sundhedsafdeling og i sidste ende til Centers for Disease Control and Prevention (CDC). Ved CDC blev stammen af gruppe A strep isoleret fra Travis analyseret mere grundigt for methicillinresistens.
Methicillinresistens er genetisk kodet og bliver mere almindelig i gruppe A strep gennem vandret genoverførsel. Ved nekrotiserende fasciitis er blodgennemstrømningen til det inficerede område typisk begrænset på grund af virkningen af forskellige genetisk kodede bakterielle toksiner., Derfor er der typisk lidt eller ingen blødning som følge af snittesten. Desværre, disse bakterielle toksiner begrænse effektiviteten af intravenøse antibiotika i clearing infektion fra huden og underliggende væv, hvilket betyder, at antibiotikaresistens alene ikke forklare ineffektivitet af Travis behandling. Ikke desto mindre var intravenøs antibiotikabehandling berettiget til at hjælpe med at minimere det mulige resultat af sepsis, hvilket er et almindeligt resultat af nekrotiserende fasciitis., Gennem genomisk analyse af CDC af den stamme, der er isoleret fra Travis, flere af de vigtige virulens-gener, der viste sig at være kodet på prophages, hvilket indikerer, at transduktion er vigtigt i horisontal gen-overførsel af disse gener fra en bakteriecelle til en anden.
nøglebegreber og resum.
- genekspression er en tæt reguleret proces.
- genekspression i prokaryoter reguleres stort set ved transkriptionspunktet. Genekspression i eukaryoter reguleres desuden post-transkriptionelt.,prokaryote strukturelle gener af relateret funktion organiseres ofte i operoner, alle kontrolleret ved transkription fra en enkelt promotor. Den regulatoriske region i en operon inkluderer promotoren selv og regionen omkring promotoren, som transkriptionsfaktorer kan binde til at påvirke transkription.
- selvom nogle operoner er konstitutivt udtrykt, er de fleste underlagt regulering ved brug af transkriptionsfaktorer (repressorer og aktivatorer)., En repressor binder til en operatør, en DNA-sekvens inden for det regulatoriske område mellem RNA-polymerasebindingsstedet i promotoren og det første strukturelle gen, hvorved fysisk blokering af transkription af disse operoner blokeres. En aktivator binder inden for det regulatoriske område af en operon, hvilket hjælper RNA-polymerase med at binde til promotoren og derved forbedre transkriptionen af denne operon. En inducer påvirker transkription gennem interaktion med en repressor eller aktivator.
- trp operon er et klassisk eksempel på en repressible operon., Når tryptophan akkumuleres, binder tryptophan sig til en repressor, som derefter binder til operatøren, hvilket forhindrer yderligere transkription.
- lac operon er et klassisk eksempel en inducerbar operon. Når lactose er til stede i cellen, omdannes den til allolactose. Allolactose virker som en inducer, der binder til undertrykkeren og forhindrer undertrykkeren i at binde til operatøren. Dette tillader transkription af de strukturelle gener.
- lac operon er også underlagt aktivering., Når glukoseniveauerne er udtømt, omdannes nogle cellulære ATP til cAMP, som binder til katabolitaktivatorproteinet (CAP). CAMP-CAP-komplekset aktiverer transkription af lac operon. Når glukoseniveauet er højt, forhindrer dets tilstedeværelse transkription af lac operon og andre operoner ved katabolitundertrykkelse.
- små intracellulære molekyler kaldet alarmones fremstilles som reaktion på forskellige miljøbelastninger, hvilket gør det muligt for bakterier at kontrollere transkriptionen af en gruppe operoner, kaldet en regulon.,
- bakterier har evnen til at ændre, hvilken RNA-faktor af RNA-polymerase de bruger som reaktion på miljøforhold for hurtigt og globalt at ændre, hvilke reguloner der transkriberes.
- prokaryoter har reguleringsmekanismer, herunder dæmpning og brug af ribosititches, til samtidig at kontrollere færdiggørelsen af transkription og oversættelse fra det transkription. Disse mekanismer arbejder gennem dannelsen af stamsløjfer i 5′ – enden af et mRNA-molekyle, der i øjeblikket syntetiseres.,
- Der er yderligere punkter til regulering af genekspression i prokaryoter og eukaryoter. I eukaryoter er epigenetisk regulering ved kemisk modifikation af DNA eller histoner og regulering af RNA-behandling to metoder.
Multiple Choice
en operon af gener, der koder for en ?ymer i en biosyntetisk vej, er sandsynligvis hvilken af følgende?
- inducible
- repressible
- konstituerende
- monocistronic
en operon-kodende gener, der transkriberes og oversættes kontinuerligt for at give cellen konstante mellemniveauer af proteinprodukterne siges at være hvilket af følgende?
- repressible
- inducible
- konstituerende
- aktiveret
hvilke af følgende betingelser fører til maksimal ekspression af lac operon?
- laktose til stede, glucose fraværende
- laktose til stede, glukose til stede
- laktose fraværende, glucose fraværende
- laktose fraværende, glukose til stede
hvilket af følgende er en type regulering af genekspression, der er unik for eukaryoter?,
- dæmpning
- brug af alternative σ faktor
- kemisk modifikation af histoner
- alarmones
udfyld det tomme
DNA-sekvensen, som undertrykkere kan binde til, der ligger mellem promotoren og det første strukturelle gen kaldes ________.,
forebyggelse af ekspression af operoner, der koder for substrat brug veje til andre substrater end glukose, når glukose er til stede kaldes _______.
tænk over det
- hvad er to måder, hvorpå bakterier kan påvirke transkriptionen af flere forskellige operoner samtidigt som reaktion på en bestemt miljøtilstand?
- følgende figur er fra Monods oprindelige arbejde med diau .isk vækst, der viser væksten af E. coli i samtidig tilstedeværelse af andylose og glucose som de eneste carbonkilder. Forklar, hvad der sker ved punkt A–D med hensyn til kulstofkilden, der bruges til vækst, og forklar, om operylose-brugsoperonen udtrykkes (og hvorfor)., Bemærk, at ekspression af de en .ymer, der kræves til xylosebrug, reguleres på en måde svarende til ekspressionen af de en .ymer, der kræves til lactosebrug.
