leerdoelstellingen
- vergelijk induceerbare operonen en repressibele operonen
- Beschrijf waarom regulatie van operonen belangrijk is
elke cel met een kern in een meercellig organisme bevat kopieën van hetzelfde DNA. Op dezelfde manier bevatten alle cellen in twee zuivere bacterieculturen geïnoculeerd uit dezelfde beginnende kolonie hetzelfde DNA, met uitzondering van veranderingen die voortvloeien uit spontane mutaties., Als elke cel in een meercellig organisme hetzelfde DNA heeft, hoe komt het dan dat cellen in verschillende delen van het lichaam van het organisme verschillende kenmerken vertonen? Evenzo, hoe komt het dat dezelfde bacteriële cellen in twee zuivere culturen blootgesteld aan verschillende omgevingsomstandigheden verschillende fenotypen kunnen vertonen? In beide gevallen zet elke genetisch identieke cel niet dezelfde set genen aan of drukt deze uit. Slechts een subset van proteã nen in een cel op een gegeven tijd wordt uitgedrukt.,
genomisch DNA bevat zowel structurele genen, die producten coderen die dienen als cellulaire structuren of enzymen, als regulerende genen, die producten coderen die genexpressie reguleren. De uitdrukking van een gen is een hoogst geregeld proces. Terwijl het regelen van genuitdrukking in multicellular organismen cellulaire differentiatie toestaat, in eencellige organismen zoals prokaryotes, verzekert het hoofdzakelijk dat de middelen van een cel niet worden verspild makend proteã nen die de cel op dat moment niet nodig heeft.,
het ophelderen van de mechanismen die de genexpressie beheersen is belangrijk voor het begrijpen van de menselijke gezondheid. Storingen in dit proces bij de mens leiden tot de ontwikkeling van kanker en andere ziekten. Het begrijpen van de interactie tussen de genexpressie van een pathogeen en die van zijn menselijke gastheer is belangrijk voor het begrijpen van een bepaalde besmettelijke ziekte. De genregelgeving impliceert een complex web van interactie binnen een bepaalde cel onder signalen van het milieu van de cel, signalerende molecules binnen de cel, en DNA van de cel., Deze interacties leiden tot de expressie van sommige genen en de onderdrukking van anderen, afhankelijk van omstandigheden.
prokaryoten en eukaryoten hebben enkele overeenkomsten in hun mechanismen om genexpressie te reguleren; echter, genexpressie in eukaryoten is ingewikkelder vanwege de tijdelijke en ruimtelijke scheiding tussen de processen van transcriptie en vertaling., Aldus, hoewel de meeste verordening van genuitdrukking door transcriptional controle in prokaryotes voorkomt, komt de verordening van genuitdrukking in eukaryotes op het transcriptional niveau en post-transcriptional voor (nadat het primaire transcript is gemaakt).
prokaryotische genregulatie
in bacteriën en archaea worden structurele eiwitten met verwante functies gewoonlijk samen gecodeerd in het genoom in een blok dat een operon wordt genoemd en worden samen getranscribeerd onder de controle van een enkele promotor, wat resulteert in de vorming van een polcistronisch transcript (figuur 1)., Op deze manier kan de regulatie van de transcriptie van alle structurele genen die coderen voor de enzymen die de vele stappen in een enkele biochemische route katalyseren, gelijktijdig worden gecontroleerd, omdat ze ofwel allemaal tegelijk nodig zijn, ofwel niet nodig zijn. Bijvoorbeeld, in E. coli, liggen alle structurele genen die enzymen coderen die nodig zijn om lactose als energiebron te gebruiken naast elkaar in het lactose (of lac) operon onder de controle van een enkele promotor, de lac promotor., De Franse wetenschappers François Jacob (1920-2013) en Jacques Monod van het Pasteur Instituut waren de eersten die de organisatie van bacteriële genen in operonen lieten zien, door hun studies naar het lac-operon van E. coli. Voor dit werk wonnen ze in 1965 de Nobelprijs voor de Fysiologie of Geneeskunde. Hoewel eukaryotic genen niet in operons worden georganiseerd, zijn prokaryotic operons uitstekende modellen voor het leren over genregelgeving in het algemeen. Er zijn sommige genclusters in eukaryotes die gelijkaardig aan operons functioneren., Veel van de principes kunnen op eukaryotic systemen worden toegepast en tot ons begrip van veranderingen in genuitdrukking in eukaryotes bijdragen die pathologische veranderingen zoals kanker kunnen resulteren.

figuur 1. In prokaryotes, worden de structurele genen van verwante functie vaak samen op het genoom georganiseerd en samen onder de controle van één enkele promotor getranscribeerd. Het regelgevend gebied van het operon omvat zowel de promotor als de exploitant. Als een onderdrukker zich bindt aan de operator, worden de structurele genen niet getranscribeerd., Als alternatief, kunnen activators aan het regelgevende gebied binden, die transcriptie verbeteren.
elk operon bevat DNA-sequenties die zijn eigen transcriptie beïnvloeden; deze bevinden zich in een gebied dat het regulerende gebied wordt genoemd. Het regelgevende gebied omvat de promotor en het gebied dat de promotor omringt, waaraan transcriptiefactoren, proteã nen die door regelgevende genen worden gecodeerd, kunnen binden. De transcriptiefactoren beà nvloeden de band van de polymerase van RNA aan de promotor en staan zijn vooruitgang toe om structurele genen te transcriberen., Een repressor is een transcriptiefactor die de transcriptie van een gen in reactie op een externe stimulus onderdrukt door aan een opeenvolging van DNA binnen het regelgevende gebied genoemd de exploitant te binden, die tussen de polymerasebindingsplaats van RNA van de promotor en de transcriptionele beginplaats van het eerste structurele gen wordt gevestigd. De band van de Repressor blokkeert fysisch RNA-polymerase van het transcriberen van structurele genen., Omgekeerd, is een activator een transcriptiefactor die de transcriptie van een gen in reactie op een externe stimulus verhoogt door de polymerase van RNA te vergemakkelijken die aan de promotor bindt. Een inductor, een derde type van regelgevende molecuul, is een kleine molecule die of transcriptie activeert of onderdrukt door in wisselwerking te staan met een repressor of een activator.
in prokaryoten zijn er voorbeelden van operonen waarvan de genproducten vrij consistent nodig zijn en waarvan de expressie daarom niet gereguleerd is., Dergelijke operons worden constitutief uitgedrukt, betekenend worden zij getranscribeerd en continu vertaald om de cel van constante tussenliggende niveaus van de eiwitproducten te voorzien. Dergelijke genen coderen enzymen betrokken bij huishoudelijke functies vereist voor cellulair onderhoud, met inbegrip van de replicatie, de reparatie, en de uitdrukking van DNA, evenals enzymen betrokken bij kernmetabolisme. In tegenstelling, zijn er andere prokaryotic operons die slechts wanneer nodig worden uitgedrukt en door onderdrukkers, activators, en inductoren worden geregeld.,
denk er eens over na
- Wat zijn de delen in de DNA-sequentie van een operon?
- welke soorten regulerende moleculen zijn er?
Regulatie door onderdrukking
prokaryotische operonen worden gewoonlijk gecontroleerd door de binding van onderdrukkers aan de operatorgebieden, waardoor transcriptie van de structurele genen wordt voorkomen. Deze operonen worden geclassificeerd als ofwel onderdrukbare operonen ofwel induceerbare operonen. Repressible operons, zoals het tryptofaan (trp) operon, bevatten typisch genen die enzymen coderen die voor een biosynthetische weg worden vereist., Zolang het product van de route, zoals tryptofaan, door de cel vereist blijft, zal een onderdrukkend operon blijven worden uitgedrukt. Nochtans, wanneer het product van de biosynthetische weg begint te accumuleren in de cel, verwijderend de behoefte aan de cel om meer te blijven maken, wordt de uitdrukking van operon onderdrukt. Omgekeerd bevatten induceerbare operonen, zoals het lac-operon van E. coli, vaak genen die enzymen coderen in een route die betrokken is bij het metabolisme van een specifiek substraat zoals lactose., Deze enzymen worden slechts vereist wanneer dat substraat beschikbaar is, aldus wordt de uitdrukking van de operons typisch veroorzaakt slechts in aanwezigheid van het substraat.
het trp-Operon: een onderdrukkend Operon
E. coli kan tryptofaan synthetiseren met behulp van enzymen die gecodeerd zijn door vijf structurele genen die naast elkaar in het trp-operon zitten (Figuur 2). Wanneer tryptofaan laag is, wordt het operon ingeschakeld. Dit betekent dat de transcriptie wordt geïnitieerd, de genen worden uitgedrukt, en tryptofaan wordt samengesteld., Als tryptofaan echter in de omgeving aanwezig is, wordt het TRP-operon uitgeschakeld. Transcriptie komt niet voor en tryptofaan wordt niet gesynthetiseerd.
wanneer tryptofaan niet in de cel aanwezig is, bindt de onderdrukker zich niet aan de operator; daarom is het operon actief en wordt tryptofaan gesynthetiseerd. Nochtans, wanneer tryptofaan in de cel accumuleert, binden twee tryptofaanmoleculen aan het TRP-onderdrukkingsmolecuul, dat zijn vorm verandert, toestaand het om aan de TRP-exploitant te binden., Deze binding van de actieve vorm van de TRP-onderdrukker aan de operator blokkeert de RNA-polymerase van het transcriberen van de structurele genen, waardoor de expressie van het operon wordt gestopt. Aldus, regelt het daadwerkelijke product van de biosynthetische weg die door het operon wordt gecontroleerd de uitdrukking van het operon.

Figuur 2. De vijf structurele genen die nodig zijn om tryptofaan in E. coli samen te stellen, bevinden zich naast elkaar in het TRP-operon. Wanneer tryptofaan afwezig is, bindt het repressoreiwit zich niet aan de operator en worden de genen getranscribeerd., Wanneer tryptofaan overvloedig aanwezig is, bindt tryptofaan het repressoreiwit aan de operatorvolgorde. Dit blokkeert fysiek de RNA-polymerase van het transcriberen van de tryptofaan biosynthesegenen.
bekijk deze video voor meer informatie over de TRP operon.
het lac-Operon: een Induceerbaar Operon
het lac-operon is een voorbeeld van een induceerbaar operon dat ook wordt geactiveerd in afwezigheid van glucose (Figuur 3)., Het lac-operon codeert voor drie structurele genen die nodig zijn om de disaccharide lactose uit het milieu te verkrijgen en te verwerken, door het op te splitsen in de eenvoudige suikers glucose en galactose. Om het lac-operon te kunnen uitdrukken, moet lactose aanwezig zijn. Dit is zinvol voor de cel omdat het energetisch verspillend zou zijn om de enzymen te creëren om lactose te verwerken als lactose niet beschikbaar was.
bij afwezigheid van lactose is de Lac-onderdrukker gebonden aan het gebied van de operator van het lac-operon, waardoor fysisch wordt voorkomen dat RNA-polymerase de structurele genen transcribeert., Echter, wanneer lactose aanwezig is, wordt de lactose in de cel omgezet in allolactose. Allolactose dient als inductormolecule, die aan de onderdrukker binden en zijn vorm veranderen zodat het niet meer aan het operatordna kan binden. De verwijdering van de repressor in aanwezigheid van lactose staat de polymerase van RNA toe om door het operatorgebied te bewegen en transcriptie van de structurele genen van lac te beginnen.
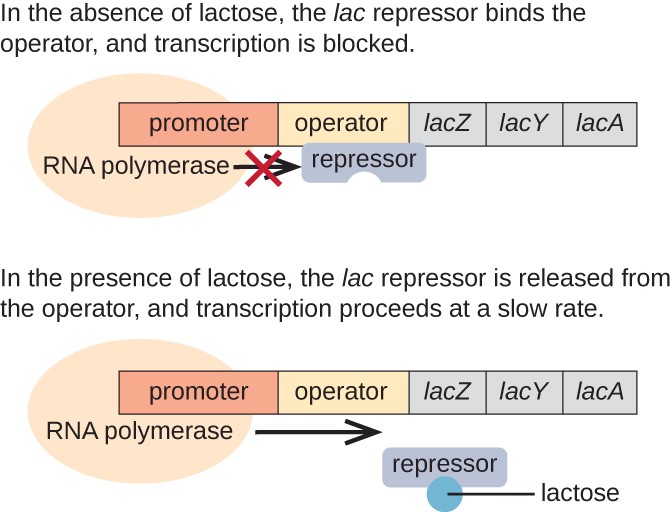
Figuur 3. De drie structurele genen die nodig zijn om lactose in E. coli te degraderen bevinden zich naast elkaar in het lac operon., Wanneer lactose afwezig is, bindt het repressoreiwit aan de operator, waardoor de RNA-polymerase fysiek wordt geblokkeerd voor het transcriberen van de Lac-structurele genen. Wanneer lactose beschikbaar is, bindt een lactosemolecuul het onderdrukkingseiwit, waardoor de onderdrukker niet aan de operatoropeenvolging bindt, en de genen worden getranscribeerd.
the lac Operon: Activation by Catabolite Activator Protein

Figuur 4. Wanneer E. coli wordt gekweekt in de aanwezigheid van twee substraten, gebruikt E. coli het gewenste substraat (in dit geval glucose) totdat het is uitgeput., Dan, enzymen nodig voor het metabolisme van het tweede substraat worden uitgedrukt en de groei hervat, hoewel aan een langzamer tarief.
bacteriën hebben doorgaans de mogelijkheid om verschillende substraten als koolstofbron te gebruiken. Omdat glucose echter meestal de voorkeur geniet boven andere substraten, hebben bacteriën mechanismen om ervoor te zorgen dat alternatieve substraten alleen worden gebruikt wanneer glucose is uitgeput. Bovendien hebben bacteriën mechanismen om ervoor te zorgen dat de genen die enzymen coderen voor het gebruik van alternatieve substraten alleen tot expressie komen wanneer het alternatieve substraat beschikbaar is., In de jaren 1940 was Jacques Monod De eerste die de voorkeur voor bepaalde substraten boven andere demonstreerde door zijn studies van E. coli ‘ s groei wanneer gecultiveerd in de aanwezigheid van twee verschillende substraten tegelijk. Dergelijke studies genereerden diauxische groeicurven, zoals in Figuur 4. Hoewel de voorkeur substraat glucose eerst wordt gebruikt, E. coli groeit snel en de enzymen voor lactose metabolisme zijn afwezig. Zodra de glucoseniveaus echter zijn uitgeput, vertragen de groeisnelheden, waardoor de expressie van de enzymen wordt veroorzaakt die nodig zijn voor het metabolisme van het tweede substraat, lactose., Merk op hoe de groeisnelheid van lactose langzamer is, zoals blijkt uit de lagere steilheid van de groeicurve.
het vermogen om van glucosegebruik over te schakelen op een ander substraat zoals lactose is een gevolg van de activiteit van een enzym genaamd enzym IIA (EIIA). Wanneer de glucosespiegels dalen, produceren cellen minder ATP van katabolisme (zie katabolisme van koolhydraten), en EIIA wordt gefosforyleerd., Phosphorylated EIIA activeert adenylylcyclase, een enzym dat sommige resterende ATP in cyclisch AMP (cAMP), een cyclisch derivaat van AMP en belangrijke signalerende molecule betrokken bij glucose en energiemetabolisme in E. coli omzet. Als gevolg daarvan, cAMP niveaus beginnen te stijgen in de cel (Figuur 5).

Figuur 5. Wanneer de ATP-niveaus wegens uitputting van glucose dalen, wordt wat resterende ATP omgezet in cAMP door adenylylcyclase. Aldus, verhoogde cAMP – niveaus signaal glucose uitputting.,
het lac-operon speelt ook een rol in deze overgang van glucose naar lactose. Wanneer glucose schaars is, bindt het accumulerende kamp dat door verhoogde adenylylcyclaseactiviteit wordt veroorzaakt aan de proteïne van de katabolietactivator (CAP), ook als proteïne van de campreceptor (CRP) wordt bekend. Het complex bindt aan het promotorgebied van het lac-operon (Figuur 6). In de regulerende gebieden van deze operonen, wordt een de verbindingsplaats van GLB gevestigd stroomopwaarts van de polymerasebindingsplaats van RNA in de promotor., De Binding van het CAP-cAMP complex aan deze plaats verhoogt de bindende capaciteit van de polymerase van RNA aan het promotorgebied om de transcriptie van de structurele genen in werking te stellen. In het geval van het lac-operon moet de transcriptie dus plaatsvinden door aanwezigheid van lactose (waarbij het Lac-onderdrukkingseiwit wordt verwijderd) en door uitputting van de glucosespiegels (waardoor binding van een activerend eiwit mogelijk wordt). Wanneer de glucoseniveaus hoog zijn, is er katabolietonderdrukking van operonen die enzymen coderen voor het metabolisme van alternatieve substraten., Vanwege de lage kampniveaus onder deze omstandigheden, is er een onvoldoende hoeveelheid van het Cap-cAMP complex om transcriptie van deze operons te activeren. Zie Tabel 1 voor een samenvatting van de regeling van het lac-operon.

Figuur 6. (A) in aanwezigheid van cAMP bindt CAP zich aan de promotors van operonen, zoals het lac-operon, die genen coderen voor enzymen voor het gebruik van alternatieve substraten. b) om het lac-operon uit te drukken, moet de Lac-onderdrukker worden geactiveerd door middel van cAMP-CAP en moet de Lac-onderdrukker van de operator worden verwijderd.,
| Tabel 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
denk er eens over na
- Wat beïnvloedt de binding van de TRP-operonrepressie aan de operator?
- Hoe en wanneer is het gedrag van het Lac-onderdrukkingseiwit veranderd?
- Hoe wordt het lac-operon niet alleen onderdrukt, maar ook gereguleerd?
globale responsen van prokaryoten
in prokaryoten zijn er ook verscheidene hogere niveaus van genregulatie die de mogelijkheid hebben om de transcriptie van vele verwante operonen gelijktijdig te controleren als reactie op een omgevingssignaal., Een groep operons die allemaal gelijktijdig bestuurd worden, wordt een regulon genoemd.
alarmsignalen
bij het waarnemen van dreigende stress veranderen prokaryoten de expressie van een grote verscheidenheid aan operonen om in coördinatie te reageren. Zij doen dit door de productie van alarmones, die kleine intracellular nucleotidederivaten zijn. Alarmones veranderen welke genen worden uitgedrukt en stimuleren de expressie van specifieke stress-respons genen. Het gebruik van alarmones om genuitdrukking in reactie op spanning te veranderen schijnt belangrijk in pathogene bacteriën te zijn., Bij het ontmoeten van gastheerverdedigingsmechanismen en andere ruwe voorwaarden tijdens besmetting, worden vele operons die virulentiegenen coderen upregulated in reactie op alarmone signaleren. Kennis van deze reacties is essentieel om het infectieproces van veel pathogenen volledig te kunnen begrijpen en om therapieën te ontwikkelen om dit proces tegen te gaan.,
alternatieve σ-factoren
aangezien de σ-subeenheid van bacterieel RNA-polymerase specificiteit geeft over welke promotors moeten worden getranscribeerd, is het wijzigen van de gebruikte σ-factor een andere manier voor bacteriën om snel en globaal te veranderen welke regulonen op een bepaald moment worden getranscribeerd. De σ-factor herkent opeenvolgingen binnen een bacteriële promotor, zodat zullen de verschillende σ-factoren elk lichtjes verschillende promotoropvolgingen erkennen., Op deze manier kan de cel, wanneer hij specifieke omgevingscondities waarneemt, reageren door de σ-factor die hij uitdrukt te veranderen, de oude te degraderen en een nieuwe te produceren om de operonen te transcriberen die genen coderen waarvan de producten nuttig zullen zijn onder de nieuwe omgevingsconditie. Bijvoorbeeld, in sporulerende bacteriën van de geslachten Bacillus en Clostridium (die vele ziekteverwekkers omvatten), controleert een groep σ factoren de uitdrukking van de vele genen die voor sporulatie in reactie op sporulation-stimulerende signalen nodig zijn.,
denk er eens over na
- Wat is de naam die wordt gegeven aan een verzameling operons die als een groep kunnen worden gereguleerd?
- welk type stimulus zou de transcriptie van een andere σ-factor veroorzaken?
aanvullende Regulatiemethoden bij bacteriën: verzwakking en Riboswitches
hoewel de meeste genexpressie wordt gereguleerd op het niveau van transcriptie-initiatie in prokaryoten, zijn er ook mechanismen om zowel de voltooiing van transcriptie als de vertaling gelijktijdig te controleren., Sinds hun ontdekking, zijn deze mechanismen getoond om de voltooiing van transcriptie en vertaling van vele prokaryotic operons te controleren. Omdat deze mechanismen de verordening van transcriptie en vertaling direct verbinden, zijn zij specifiek voor prokaryotes, omdat deze processen fysiek in eukaryotes worden gescheiden.
een dergelijk regulerend systeem is demping, waarbij secundaire stam-lusstructuren gevormd binnen het 5 ‘ einde van een mRNA die wordt getranscribeerd bepalen of transcriptie om de synthese van dit mRNA te voltooien zal plaatsvinden en of dit mRNA zal worden gebruikt voor vertaling., Naast het reeds besproken transcriptionele onderdrukkingsmechanisme controleert de verzwakking ook de expressie van het trp-operon in E. coli (Figuur 7). Het TRP-operonreguleringsgebied bevat een leader-sequentie genaamd trpL tussen de operator en het eerste structurele gen, dat vier stukken RNA heeft die paar met elkaar in verschillende combinaties kunnen baseren. Wanneer een terminatorstam-lijn vormt, beëindigt de transcriptie, vrijgevend de polymerase van RNA van mRNA., Nochtans, wanneer een antiterminator stam-lijn vormt, verhindert dit de vorming van de terminator stam-lijn, zodat kan de polymerase van RNA de structurele genen transcriberen.
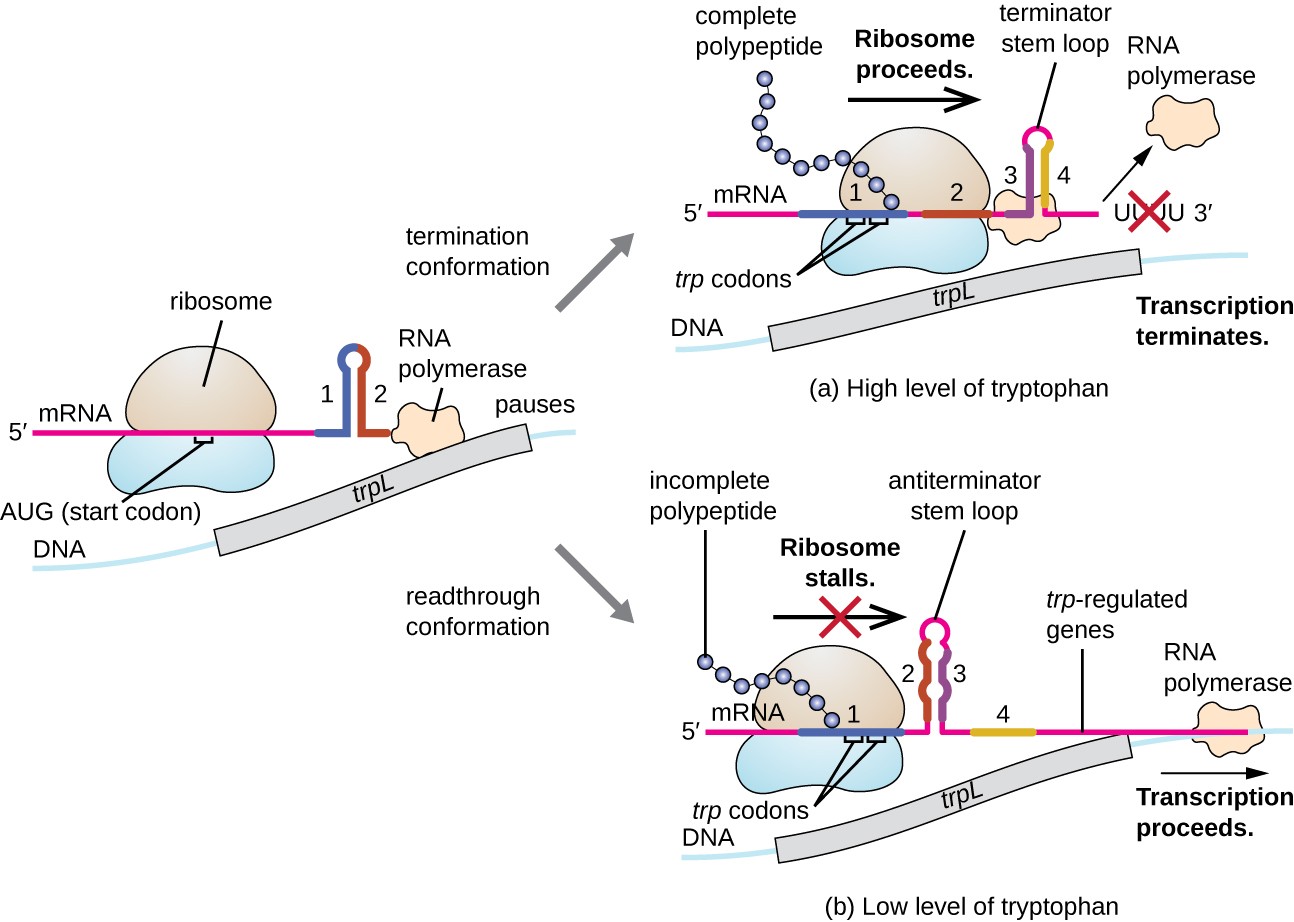
Figuur 7. Klik om een grotere afbeelding te bekijken. Wanneer tryptofaan overvloedig is, verloopt de vertaling van het korte leader peptide dat door trpL wordt gecodeerd, De terminatorlus tussen gebieden 3 en 4 vormen, en de transcriptie eindigt., Wanneer de tryptofaanniveaus uitgeput zijn, kraamt de vertaling van de korte leader-peptide op Gebied 1, die gebieden 2 en 3 toestaat om een antiterminatorlus te vormen, en de polymerase van RNA kan de structurele genen van het TRP-operon transcriberen.
een verwant mechanisme van gelijktijdige regulatie van transcriptie en vertaling in prokaryoten is het gebruik van een riboswitch, een klein gebied van noncoding RNA dat wordt gevonden binnen het 5′ – einde van sommige prokaryotische mRNA-moleculen (Figuur 8). Een riboswitch kan aan een kleine intracellular molecule binden om bepaalde secundaire structuren van de mRNA-molecule te stabiliseren., De band van de kleine molecule bepaalt welke de structuur van de stam-lijn vormt, waarbij de voltooiing van mRNA-synthese en eiwitsynthese wordt beà nvloed.

Figuur 8. Klik voor een grotere afbeelding. De riboswitches die binnen prokaryotic mRNA molecules worden gevonden kunnen aan kleine intracellular molecules binden, die bepaalde structuren van RNA stabiliseren, of de voltooiing van de synthese van de molecule van mRNA zelf (links) beà nvloeden of de proteã ne die wordt gemaakt gebruikend dat mRNA (rechts).,
andere factoren die de genexpressie in prokaryoten en eukaryoten beïnvloeden
hoewel de focus op onze discussie over transcriptionele controle prokaryotische operonen als voorbeelden gebruikte, is eukaryotische transcriptionele controle in vele opzichten vergelijkbaar. Zoals in prokaryotes, kan de eukaryotic transcriptie door de band van transcriptiefactoren met inbegrip van onderdrukkers en activators worden gecontroleerd., Interessant, kan de eukaryotic transcriptie door de band van proteã nen aan gebieden van DNA worden beà nvloed, genoemd versterkers, vrij ver van het gen, door het van een lus voorzien van DNA tussen de versterker en de promotor (figuur 9). Globaal, is het regelen van transcriptie een hoogst efficiënte manier om genuitdrukking in zowel prokaryotes als eukaryotes te controleren. Nochtans, kan de controle van genuitdrukking in eukaryotes in reactie op milieu en cellulaire spanningen op extra manieren zonder de band van transcriptiefactoren aan regelgevende gebieden worden bereikt.,

figuur 9. In eukaryotes, is een versterker een opeenvolging van DNA die transcriptie bevordert. Elke versterker bestaat uit korte DNA-sequenties die distale controle-elementen worden genoemd. De aan de distale controle-elementen gebonden activatoren interageren met mediatorproteã nen en transcriptiefactoren. Twee verschillende genen kunnen dezelfde promotor hebben maar verschillende distale controle-elementen, waardoor differentiële genexpressie mogelijk is.,
controle op het DNA-niveau
In eukaryoten kunnen de DNA-moleculen of geassocieerde histonen chemisch worden gewijzigd om de transcriptie te beïnvloeden; dit wordt epigenetische Regulatie genoemd. Methylation van bepaalde cytosinenucleotiden in DNA in reactie op milieufactoren is getoond om gebruik van dergelijk DNA voor transcriptie te beà nvloeden, met methylation van DNA algemeen correlerend aan verlaagde niveaus van genuitdrukking., Bovendien, in reactie op milieufactoren, kunnen histone proteã nen voor verpakking DNA ook Chemisch op veelvoudige manieren, met inbegrip van acetylation en deacetylation worden gewijzigd, beà nvloedend de verpakkingstoestand van DNA en zo beà nvloedend de beschikbaarheid van losjes gekronkelde DNA voor transcriptie. Deze chemische wijzigingen kunnen soms door veelvoudige rondes van celdeling worden gehandhaafd, die minstens sommige van deze epigenetische veranderingen erfelijk maken.
deze video beschrijft hoe epigenetische Regulatie de genexpressie controleert.,
denk er eens over na
- Wat stopt of staat transcriptie toe om door te gaan als demping werkt?
- Wat bepaalt de status van een riboswitch?
- Beschrijf de functie van een versterker.
- Beschrijf twee mechanismen van epigenetische Regulatie in eukaryoten.
klinische Focus: Travis, resolutie
Dit voorbeeld concludeert Travis ‘ verhaal dat begon in de functies van genetisch materiaal, RNA-transcriptie, en hoe aseksuele prokaryoten genetische diversiteit bereiken.,hoewel Travis zijn aanval met necrotiserende fasciitis overleefde, moest hij nu een huidtransplantatie ondergaan, gevolgd door langdurige fysiotherapie. Op basis van de hoeveelheid spiermassa die hij verloor, is het onwaarschijnlijk dat zijn been op volle kracht zal terugkeren, maar zijn fysiotherapeut is optimistisch dat hij zijn been weer zal gebruiken.uit laboratoriumonderzoek bleek dat de veroorzaker van Travis ‘ infectie een stam was van streptococcus groep A (streptococcus groep A)., Zoals vereist door de wet, Travis ‘ s zaak werd gemeld aan de State Health department en uiteindelijk aan de Centers for Disease Control and Prevention (CDC). Bij het CDC werd de stam van groep a strep geïsoleerd van Travis grondiger geanalyseerd op methicilline resistentie.
resistentie tegen methicilline is genetisch gecodeerd en komt vaker voor in groep a strep via horizontale genoverdracht. In necrotiserende fasciitis, is de bloedstroom naar het besmette gebied typisch beperkt wegens de actie van diverse genetisch gecodeerde bacteriële toxines., Dit is de reden waarom er meestal weinig tot geen bloeden als gevolg van de incisie test. Helaas beperken deze bacteriële toxines de effectiviteit van intraveneuze antibiotica bij het verwijderen van infecties van de huid en het onderliggende weefsel, wat betekent dat antibioticaresistentie alleen niet de ineffectiviteit van de behandeling van Travis verklaart. Niettemin, intraveneuze antibiotische therapie werd gerechtvaardigd om het mogelijke resultaat van sepsis te helpen minimaliseren, die een gemeenschappelijk resultaat van necrotiserende fasciitis is., Door genomic analyse door CDC van de stam die van Travis wordt geà soleerd, werden verscheidene belangrijke virulentiegenen getoond om op profages worden gecodeerd, die erop wijzen dat transductie in de horizontale genoverdracht van deze genen van één bacteriële cel aan een andere belangrijk is.
sleutelconcepten en samenvatting
- genexpressie is een strak gereguleerd proces.
- genexpressie in prokaryoten wordt grotendeels geregeld op het punt van transcriptie. De genuitdrukking in eukaryotes is bovendien post-transcriptioneel geregeld.,
- prokaryotische structurele genen met verwante functie worden vaak georganiseerd in operonen, alle gecontroleerd door transcriptie van een enkele promotor. Het regelgevende gebied van een operon omvat de promotor zelf en het gebied dat de promotor omringt waaraan transcriptiefactoren kunnen binden om transcriptie te beïnvloeden.
- hoewel sommige operonen constitutief worden uitgedrukt, zijn de meeste onderworpen aan Regulatie door het gebruik van transcriptiefactoren (onderdrukkers en activatoren)., Een repressor bindt aan een operator, een DNA-sequentie binnen het regulerende gebied tussen de RNA-polymerasebindingsplaats in de promotor en het eerste structurele gen, waardoor de transcriptie van deze operonen fysiek wordt geblokkeerd. Een activator bindt binnen het regelgevende gebied van een operon, helpend de polymerase van RNA bindt aan de promotor, daardoor verbeterend de transcriptie van dit operon. Een inductor beïnvloedt de transcriptie door interactie met een onderdrukker of activator.
- het TRP-operon is een klassiek voorbeeld van een onderdrukbare operon., Wanneer tryptofaan accumuleert, bindt tryptofaan aan een onderdrukker, die dan aan de exploitant bindt, die verdere transcriptie verhindert.
- het lac-operon is een klassiek voorbeeld van een induceerbare operon. Wanneer lactose in de cel aanwezig is, wordt het omgezet in allolactose. Allolactose werkt als inductor, bindt aan de onderdrukker en voorkomt dat de onderdrukker bindt aan de operator. Dit staat transcriptie van de structurele genen toe.
- het lac-operon wordt ook geactiveerd., Wanneer de glucoseniveaus uitgeput zijn, wordt wat cellulair ATP omgezet in kamp, dat aan de katabolite activatorproteã ne (GLB) bindt. Het cAMP-CAP complex activeert de transcriptie van het lac operon. Wanneer de glucoseniveaus hoog zijn, verhindert zijn aanwezigheid transcriptie van het lac-operon en andere operons door katabolietonderdrukking.
- kleine intracellulaire moleculen, alarmones genoemd, worden gemaakt als reactie op verschillende omgevingsstress, waardoor bacteriën de transcriptie van een groep operonen, een regulon genaamd, kunnen controleren.,
- bacteriën hebben het vermogen om te veranderen welke σ-factor van RNA-polymerase zij gebruiken als reactie op omgevingsomstandigheden om snel en globaal te veranderen welke regulonen worden getranscribeerd.
- prokaryoten hebben regulerende mechanismen, waaronder verzwakking en het gebruik van riboswitches, om tegelijkertijd de voltooiing van transcriptie en vertaling van dat transcript te controleren. Deze mechanismen werken door de vorming van stamlussen in het einde 5′ van een mRNA-molecule die momenteel wordt samengesteld.,
- Er zijn extra regulatiepunten van genexpressie in prokaryoten en eukaryoten. In eukaryotes, zijn de epigenetische regelgeving door chemische wijziging van DNA of histones, en de regelgeving van de verwerking van RNA twee methodes.
Multiple Choice
een operon van genen die enzymen coderen in een biosynthetische route is waarschijnlijk welke van de volgende?
- inducible
- repressible
- constitutive
- monocistronic
een operon dat genen codeert die continu worden getranscribeerd en vertaald om de cel te voorzien van constante intermediaire niveaus van de eiwitproducten, zou zijn welke van de volgende?
- repressible
- inducible
- constitutief
- geactiveerd
welke van de volgende voorwaarden leidt tot maximale expressie van het lac-operon?
- lactose aanwezig is, glucose afwezig
- lactose aanwezig is, glucose aanwezig
- lactose afwezig, glucose afwezig
- lactose afwezig, glucose aanwezig
welke van de volgende is een soort regulatie van genexpressie uniek voor eukaryoten?,
- verzwakking
- gebruik van alternatieve σ factor
- chemische modificatie van histonen
- alarmonen
vul de blanco in
de DNA-sequentie die tussen de promotor en het eerste structurele gen ligt, wordt ________ _ genoemd.,
de preventie van expressie van operonen die coderen voor substraat gebruik routes voor andere substraten dan glucose wanneer glucose aanwezig is wordt _______genoemd.
denk er eens over na
- Wat zijn twee manieren waarop bacteriën de transcriptie van meerdere verschillende operonen gelijktijdig kunnen beïnvloeden in reactie op een bepaalde milieuconditie?
- het volgende cijfer is afkomstig van Monod ‘ s oorspronkelijke werk over diauxische groei, waaruit de groei van E. coli blijkt in de gelijktijdige aanwezigheid van xylose en glucose als de enige koolstofbronnen. Leg uit wat er gebeurt op de punten A–D met betrekking tot de koolstofbron die wordt gebruikt voor de groei, en leg uit of het xylose-gebruik operon wordt uitgedrukt (en waarom)., Merk op dat de expressie van de enzymen die nodig zijn voor het gebruik van xylose wordt geregeld op een manier die vergelijkbaar is met de expressie van de enzymen die nodig zijn voor het gebruik van lactose.
