cele uczenia się
- Porównaj indukcyjne operony i represyjne operony
- opisz dlaczego Regulacja operonów jest ważna
każda komórka nukleacyjna w wielokomórkowym organizmie zawiera kopie tego samego DNA. Podobnie, wszystkie komórki w dwóch czystych kulturach bakteryjnych zaszczepionych z tej samej Kolonii wyjściowej zawierają to samo DNA, z wyjątkiem zmian, które powstają w wyniku spontanicznych mutacji., Jeśli każda komórka w organizmie wielokomórkowym ma to samo DNA, to jak to jest, że komórki w różnych częściach ciała organizmu wykazują różne cechy? Podobnie, jak to jest, że te same komórki bakteryjne w dwóch czystych kulturach narażonych na różne warunki środowiskowe mogą wykazywać różne fenotypy? W obu przypadkach każda genetycznie identyczna komórka nie włącza ani nie wyraża tego samego zestawu genów. Wyraża się tylko podzbiór białek w komórce w danym czasie.,
genomowe DNA zawiera zarówno geny strukturalne, które kodują produkty, które służą jako struktury komórkowe lub enzymy, jak i geny regulacyjne, które kodują produkty, które regulują ekspresję genów. Ekspresja genu jest procesem wysoce regulowanym. Podczas gdy regulowanie ekspresji genów w organizmach wielokomórkowych pozwala na różnicowanie komórkowe, w organizmach jednokomórkowych, takich jak prokarioty, to przede wszystkim zapewnia, że zasoby komórki nie są marnowane tworząc białka, które komórka nie potrzebuje w tym czasie.,
Wyjaśnienie mechanizmów kontrolujących ekspresję genów jest ważne dla zrozumienia ludzkiego zdrowia. Nieprawidłowości w tym procesie u ludzi prowadzą do rozwoju raka i innych chorób. Zrozumienie interakcji między ekspresją genu patogenu a ekspresją jego ludzkiego gospodarza jest ważne dla zrozumienia konkretnej choroby zakaźnej. Regulacja genów obejmuje złożoną sieć interakcji w obrębie danej komórki między sygnałami ze środowiska komórki, cząsteczkami sygnalizacyjnymi w komórce i DNA komórki., Interakcje te prowadzą do ekspresji niektórych genów i tłumienia innych, w zależności od okoliczności.
Prokaryotes i eukaryotes dzielić niektóre podobieństwa w ich mechanizmach regulować gene ekspresję; jakkolwiek, gene ekspresja w eukaryotes jest bardziej skomplikowany ponieważ czasowy i przestrzenny rozdzielenie między procesami transkrypcja i translacja., Tak więc, chociaż większość regulacji ekspresji genów występuje poprzez kontrolę transkrypcyjną u prokariotów, regulacja ekspresji genów u eukariotów występuje na poziomie transkrypcyjnym i post-transkrypcyjnym (po pierwotnym transkrypcie został wykonany).
Prokariotyczna Regulacja genu
u bakterii i archeonów białka strukturalne o powiązanych funkcjach są zwykle kodowane razem w genomie w bloku zwanym OPERONEM i są transkrybowane razem pod kontrolą pojedynczego promotora, co powoduje powstanie transkrypcji polihistronicznej (ryc. 1)., W ten sposób Regulacja transkrypcji wszystkich genów strukturalnych kodujących enzymy katalizujące wiele etapów w jednym szlaku biochemicznym może być kontrolowana jednocześnie, ponieważ albo wszystkie będą potrzebne w tym samym czasie, albo żaden nie będzie potrzebny. Na przykład w E. coli wszystkie geny strukturalne kodujące enzymy potrzebne do wykorzystania laktozy jako źródła energii leżą obok siebie w operonie laktozy (lub lac) pod kontrolą jednego promotora, promotora lac., Francuscy naukowcy François Jacob (1920-2013) i Jacques Monod z Instytutu Pasteura jako pierwsi wykazali organizację genów bakteryjnych w operonach poprzez badania nad lac OPERONEM E. coli. Za tę pracę otrzymał Nagrodę Nobla w dziedzinie fizjologii lub medycyny w 1965 roku. Chociaż geny eukariotyczne nie są zorganizowane w operony, prokariotyczne operony są doskonałymi modelami do nauki o regulacji genów ogólnie. Istnieją pewne klastry genów u eukariotów, które funkcjonują podobnie do operonów., Wiele z zasad może być stosowane do systemów eukariotycznych i przyczynić się do naszego zrozumienia zmian w ekspresji genów u eukariotów, które mogą powodować zmiany patologiczne, takie jak rak.

Rysunek 1. U prokariotów, strukturalne geny pokrewnych funkcji są często organizowane razem na genomie i transkrybowane razem pod kontrolą jednego promotora. Region regulacyjny Operonu obejmuje zarówno organizatora, jak i operatora. Jeśli represor wiąże się z operatorem, geny strukturalne nie zostaną transkrybowane., Alternatywnie aktywatory mogą wiązać się z regionem regulacyjnym, zwiększając transkrypcję.
każdy operon zawiera sekwencje DNA, które wpływają na jego własną transkrypcję; znajdują się one w regionie zwanym regionem regulacyjnym. Region regulacyjny obejmuje promotor i region otaczający promotor, z którym mogą wiązać się czynniki transkrypcyjne, białka kodowane przez geny regulatorowe. Czynniki transkrypcyjne wpływają na Wiązanie polimerazy RNA z promotorem i pozwalają jej progresji na transkrypcję genów strukturalnych., Represor jest czynnikiem transkrypcyjnym, który hamuje transkrypcję genu w odpowiedzi na bodziec zewnętrzny poprzez wiązanie się z sekwencją DNA w obrębie regionu regulacyjnego zwanego operatorem, który znajduje się między miejscem wiązania polimerazy RNA promotora i miejscem rozpoczęcia transkrypcji pierwszego strukturalnego genu. Wiązanie represyjne fizycznie blokuje polimerazę RNA przed transkrypcją genów strukturalnych., Natomiast aktywator jest czynnikiem transkrypcyjnym, który zwiększa transkrypcję genu w odpowiedzi na bodziec zewnętrzny, ułatwiając Wiązanie polimerazy RNA z promotorem. Induktor, trzeci typ cząsteczki regulacyjnej, jest małą cząsteczką, która aktywuje lub hamuje transkrypcję poprzez interakcję z represorem lub aktywatorem.
u prokariotów istnieją przykłady operonów, których produkty genowe są wymagane dość konsekwentnie i których ekspresja jest zatem nieuregulowana., Takie operony są konstytutywnie wyrażone, co oznacza, że są transkrybowane i tłumaczone w sposób ciągły, aby zapewnić komórce stały pośredni poziom produktów białkowych. Geny te kodują enzymy zaangażowane w funkcje utrzymania komórek, w tym replikację, naprawę i ekspresję DNA, a także enzymy zaangażowane w metabolizm rdzenia. W przeciwieństwie do tego, istnieją inne prokariotyczne operony, które są wyrażane tylko wtedy, gdy są potrzebne i są regulowane przez represory, aktywatory i induktory.,
pomyśl o tym
- jakie są części w sekwencji DNA Operonu?
- jakie są rodzaje cząsteczek regulacyjnych?
Regulacja przez represje
Operony prokariotyczne są powszechnie kontrolowane przez wiązanie represji do regionów operatora, zapobiegając w ten sposób transkrypcji genów strukturalnych. Takie operony są klasyfikowane jako operony represyjne lub indukcyjne. Operony represyjne, podobnie jak tryptofan (trp) operon, zazwyczaj zawierają geny kodujące enzymy wymagane dla szlaku biosyntetycznego., Tak długo, jak produkt szlaku, jak tryptofan, nadal jest wymagany przez komórkę, represywny operon będzie nadal wyrażany. Jednak, gdy produkt szlaku biosyntetycznego zaczyna gromadzić się w komórce, eliminując potrzebę, aby komórka nadal wytwarzała więcej, ekspresja Operonu jest tłumiona. Z drugiej strony, indukowane operony, jak lac operon E. coli, często zawierają geny kodujące enzymy w szlaku metabolicznym zaangażowanym w metabolizm określonego substratu, takiego jak laktoza., Enzymy te są wymagane tylko wtedy, gdy substrat jest dostępny, a zatem ekspresja operonów jest zwykle indukowana tylko w obecności substratu.
trp Operon: represyjny Operon
E. coli może syntetyzować tryptofan za pomocą enzymów, które są kodowane przez pięć genów strukturalnych znajdujących się obok siebie w trp operon (ryc. 2). Gdy tryptofan środowiskowy jest niski, operon jest włączony. Oznacza to, że transkrypcja jest inicjowana, geny są wyrażane, a tryptofan jest syntetyzowany., Jednak jeśli tryptofan jest obecny w środowisku, trp operon jest wyłączony. Transkrypcja nie zachodzi, a tryptofan nie jest syntetyzowany.
gdy tryptofan nie jest obecny w komórce, represor sam w sobie nie wiąże się z operatorem, dlatego operon jest aktywny i tryptofan jest syntetyzowany. Jednak gdy tryptofan gromadzi się w komórce, dwie cząsteczki tryptofanu wiążą się z cząsteczką represora trp, która zmienia swój kształt, umożliwiając Wiązanie się z operatorem trp., Wiązanie aktywnej formy represora trp z operatorem blokuje polimerazę RNA przed transkrypcją genów strukturalnych, zatrzymując ekspresję Operonu. Tak więc rzeczywisty produkt szlaku biosyntetycznego kontrolowanego przez operon reguluje ekspresję Operonu.

Rysunek 2. Pięć genów strukturalnych potrzebnych do syntezy tryptofanu w E. coli znajduje się obok siebie w trp operon. Gdy tryptofan jest nieobecny, białko represora nie wiąże się z operatorem, a geny są transkrybowane., Gdy tryptofan jest obfity, tryptofan wiąże białko represor w sekwencji operatora. To fizycznie blokuje polimerazę RNA przed transkrypcją genów biosyntezy tryptofanu.
obejrzyj ten film, aby dowiedzieć się więcej o trp operon.
lac Operon: indukowany Operon
Lac operon jest przykładem indukowanego Operonu, który również podlega aktywacji w przypadku braku glukozy (ryc. 3)., Lac operon koduje trzy geny strukturalne niezbędne do pozyskiwania i przetwarzania laktozy disacharydowej z otoczenia, rozkładając ją na cukry proste-glukozę i galaktozę. Aby lac operon mógł być wyrażony, musi być obecna laktoza. Ma to sens dla komórki, ponieważ byłoby energicznie marnotrawne tworzenie enzymów do przetwarzania laktozy, gdyby laktoza nie była dostępna.
w przypadku braku laktozy represor lac wiąże się z regionem operatora lac Operonu, fizycznie uniemożliwiając polimerazie RNA transkrypcję genów strukturalnych., Jednakże, gdy laktoza jest obecna, laktoza wewnątrz komórki jest przekształcana w allolaktozę. Allolaktoza służy jako cząsteczka indukująca, wiążąca się z represorem i zmieniająca swój kształt tak, że nie jest już w stanie wiązać się z DNA operatora. Usunięcie represora w obecności laktozy umożliwia polimerazie RNA przemieszczanie się przez region operatora i rozpoczęcie transkrypcji genów strukturalnych lac.
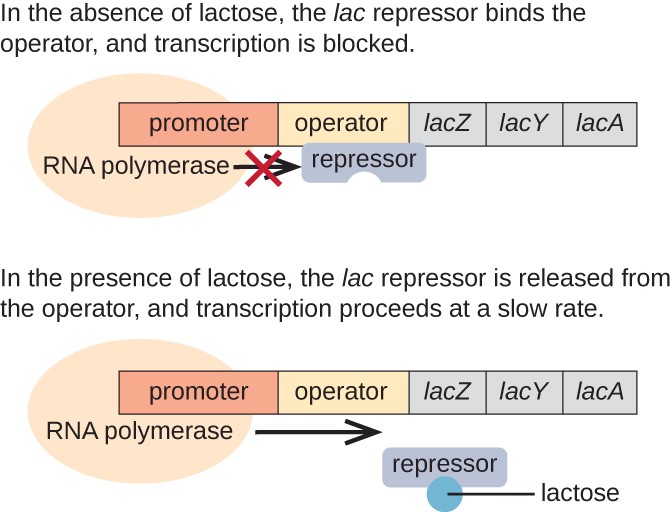
Rysunek 3. Trzy geny strukturalne, które są potrzebne do degradacji laktozy w E. coli, znajdują się obok siebie w lac operon., Gdy laktoza jest nieobecna, białko represyjne wiąże się z operatorem, fizycznie blokując polimerazę RNA przed transkrypcją genów lac. Gdy laktoza jest dostępna, cząsteczka laktozy wiąże białko represyjne, uniemożliwiając Wiązanie represyjne z sekwencją operatora, a geny są transkrybowane.
Lac Operon: aktywacja przez białko aktywatora Katabolitu

Rysunek 4. W przypadku uprawy w obecności dwóch substratów, E. coli używa preferowanego substratu (w tym przypadku glukozy), dopóki nie zostanie wyczerpany., Następnie enzymy potrzebne do metabolizmu drugiego substratu są wyrażane i wzrost wznawia się, choć w wolniejszym tempie.
Jednakże, ponieważ glukoza jest zwykle korzystniejsza od innych substratów, bakterie mają mechanizmy zapewniające, że alternatywne substraty są używane tylko wtedy, gdy glukoza została zubożona. Dodatkowo bakterie mają mechanizmy zapewniające, że geny kodujące enzymy do stosowania substratów alternatywnych są wyrażane tylko wtedy, gdy substrat alternatywny jest dostępny., W latach 40. Jacques Monod jako pierwszy zademonstrował preferencję niektórych substratów w stosunku do innych, badając wzrost bakterii E. coli podczas hodowli w obecności dwóch różnych substratów jednocześnie. Takie badania wygenerowały diauxic krzywe wzrostu, jak ten pokazany na rysunku 4. Chociaż preferowanym substratem jest glukoza, E. coli szybko rośnie, a enzymy metabolizmu laktozy są nieobecne. Jednakże, gdy poziom glukozy zostanie wyczerpany, tempo wzrostu spowolni się, indukując ekspresję enzymów potrzebnych do metabolizmu drugiego substratu, laktozy., Zauważ, że tempo wzrostu laktozy jest wolniejsze, na co wskazuje niższa stromo krzywa wzrostu.
zdolność do przejścia z glukozy na inny substrat, taki jak laktoza, jest konsekwencją aktywności enzymu zwanego enzymem IIA (EIIA). Gdy poziom glukozy spada, komórki produkują mniej ATP z katabolizmu (zobacz katabolizm węglowodanów), a EIIA ulega fosforylacji., Fosforylowana EIIA aktywuje cyklazę adenylową, enzym, który przekształca część pozostałego ATP w cykliczny AMP (cAMP), cykliczną pochodną AMP i ważną cząsteczkę sygnalizacyjną zaangażowaną w metabolizm glukozy i energii w E. coli. W rezultacie poziom obozu zaczyna wzrastać w komórce (ryc. 5).

Rysunek 5. Gdy poziom ATP zmniejsza się z powodu wyczerpania glukozy, część pozostała ATP jest przekształcana w Camp przez cyklazę adenylową. W ten sposób zwiększony poziom cAMP sygnalizuje wyczerpanie glukozy.,
lac operon odgrywa również rolę w tym przełączeniu z glukozy na laktozę. Gdy glukoza jest rzadka, akumulujący cAMP spowodowany zwiększoną aktywnością cyklazy adenylowej wiąże się z białkiem aktywatora katabolitu (CAP), znanym również jako białko receptora cAMP (CRP). Kompleks wiąże się z regionem promotora lac operon (ryc. 6). W regionach regulacyjnych tych operonów miejsce wiązania Kapu znajduje się przed miejscem wiązania polimerazy RNA w promotorze., Wiązanie kompleksu CAP-cAMP z tym miejscem zwiększa zdolność wiązania polimerazy RNA z regionem promotora do inicjowania transkrypcji genów strukturalnych. Tak więc w przypadku Operonu lac, aby doszło do transkrypcji, musi być obecna laktoza (usunięcie białka Lac), a poziom glukozy musi być obniżony(umożliwienie wiązania białka aktywującego). Gdy poziom glukozy jest wysoki, dochodzi do katabolizmu operonów kodujących enzymy dla metabolizmu substratów alternatywnych., Ze względu na niski poziom obozu w tych warunkach, nie ma wystarczającej ilości kompleksu CAP-cAMP, aby aktywować transkrypcję tych operonów. Streszczenie regulacji lac operon znajduje się w tabeli 1.

Rysunek 6. a) w obecności cAMP, CAP wiąże się z promotorami operonów, takimi jak lac operon, które kodują geny dla enzymów do stosowania substratów alternatywnych. b) dla wyrażenia Operonu lac musi być aktywacja przez Camp-CAP, jak również usunięcie tłumika lac od operatora.,
| Tabela 1.,ion of the lac Operon | ||||
|---|---|---|---|---|
| Glucose | CAP binds | Lactose | Repressor binds | Transcription |
| + | – | – | + | No |
| + | – | + | – | Some |
| – | + | – | + | No |
| – | + | + | – | Yes |
Watch an animated tutorial about the workings of lac operon here.,
pomyśl o tym
- co wpływa na Wiązanie Operonu trp z operatorem?
- jak i kiedy zmienia się zachowanie białka lac?
- poza tym, że jest represyjny, jak inaczej jest regulowany lac operon?
Globalne reakcje prokariotów
w prokariotach istnieje również kilka wyższych poziomów regulacji genów, które mają zdolność kontrolowania transkrypcji wielu powiązanych operonów jednocześnie w odpowiedzi na sygnał środowiskowy., Grupa operonów sterowanych jednocześnie nazywana jest regulonem.
Alarmony
podczas wyczuwania zbliżającego się stresu prokaryoty zmieniają ekspresję wielu różnych operonów, aby reagować w koordynacji. Robią to poprzez produkcję alarmonów, które są małymi wewnątrzkomórkowymi pochodnymi nukleotydów. Alarmony zmieniają, które geny są wyrażane i stymulują ekspresję specyficznych genów odpowiedzi stresowej. Zastosowanie alarmonów do zmiany ekspresji genów w odpowiedzi na stres wydaje się być ważne w chorobotwórczych bakteriach., Po napotkaniu mechanizmów obronnych gospodarza i innych trudnych warunków podczas infekcji, wiele operonów kodujących geny zjadliwości jest regulowanych w odpowiedzi na sygnalizację alarmową. Znajomość tych reakcji jest kluczem do bycia w stanie w pełni zrozumieć proces infekcji wielu patogenów i rozwoju terapii przeciwdziałających temu procesowi.,
alternatywne σ czynniki
ponieważ σ podjednostka bakteryjnej polimerazy RNA nadaje swoistość co do tego, które promotory powinny być transkrybowane, zmiana zastosowanego σ czynnika jest innym sposobem dla bakterii, aby szybko i globalnie zmienić to, co regulony są transkrybowane w danym czasie. Czynnik σ rozpoznaje sekwencje wewnątrz promotora bakteryjnego, więc różne czynniki σ rozpoznają nieco inne sekwencje promotora., W ten sposób, gdy komórka wyczuwa określone warunki środowiskowe, może reagować poprzez zmianę wyrażonego przez nią czynnika σ, degradując stary i produkując Nowy, aby przepisać operony kodujące geny, których produkty będą przydatne w nowym stanie środowiska. Na przykład u bakterii sporulujących z rodzajów Bacillus i Clostridium (które zawierają wiele patogenów), grupa czynników σ kontroluje ekspresję wielu genów potrzebnych do sporulacji w odpowiedzi na sygnały stymulujące sporulację.,
pomyśl o tym
- Jak nazywa się zbiór operonów, które można regulować jako grupę?
- jaki rodzaj bodźca wywołałby transkrypcję innego czynnika σ?
dodatkowe metody regulacji u bakterii: tłumienie i Ryboswitche
chociaż większość ekspresji genów jest regulowana na poziomie inicjacji transkrypcji u prokariotów, istnieją również mechanizmy kontroli zarówno zakończenia transkrypcji, jak i translacji jednocześnie., Od czasu ich odkrycia wykazano, że mechanizmy te kontrolują zakończenie transkrypcji i translacji wielu prokariotycznych operonów. Ponieważ mechanizmy te łączą regulację transkrypcji i translacji bezpośrednio, są one specyficzne dla prokaryotes, ponieważ procesy te są fizycznie oddzielone w eukaryotes.
jednym z takich systemów regulacyjnych jest tłumienie, w którym wtórne struktury pętli macierzystej utworzone w 5′ końcu transkrypcji mRNA określają, czy nastąpi transkrypcja w celu zakończenia syntezy tego mRNA i czy ten mRNA zostanie użyty do tłumaczenia., Poza omówionym już mechanizmem represji transkrypcyjnej, tłumienie kontroluje również ekspresję trp operon u E. coli (ryc. 7). Region regulacyjny trp operon zawiera sekwencję leader zwaną trpL pomiędzy operatorem a pierwszym genem strukturalnym, który ma cztery odcinki RNA, które mogą bazować ze sobą w różnych kombinacjach. Kiedy tworzy się pętla macierzysta terminatora, transkrypcja kończy się, uwalniając polimerazę RNA z mRNA., Jednak, gdy tworzy się antytererminatorowa pętla macierzysta, zapobiega to tworzeniu się terminatorowej pętli macierzystej, więc polimeraza RNA może przepisać geny strukturalne.
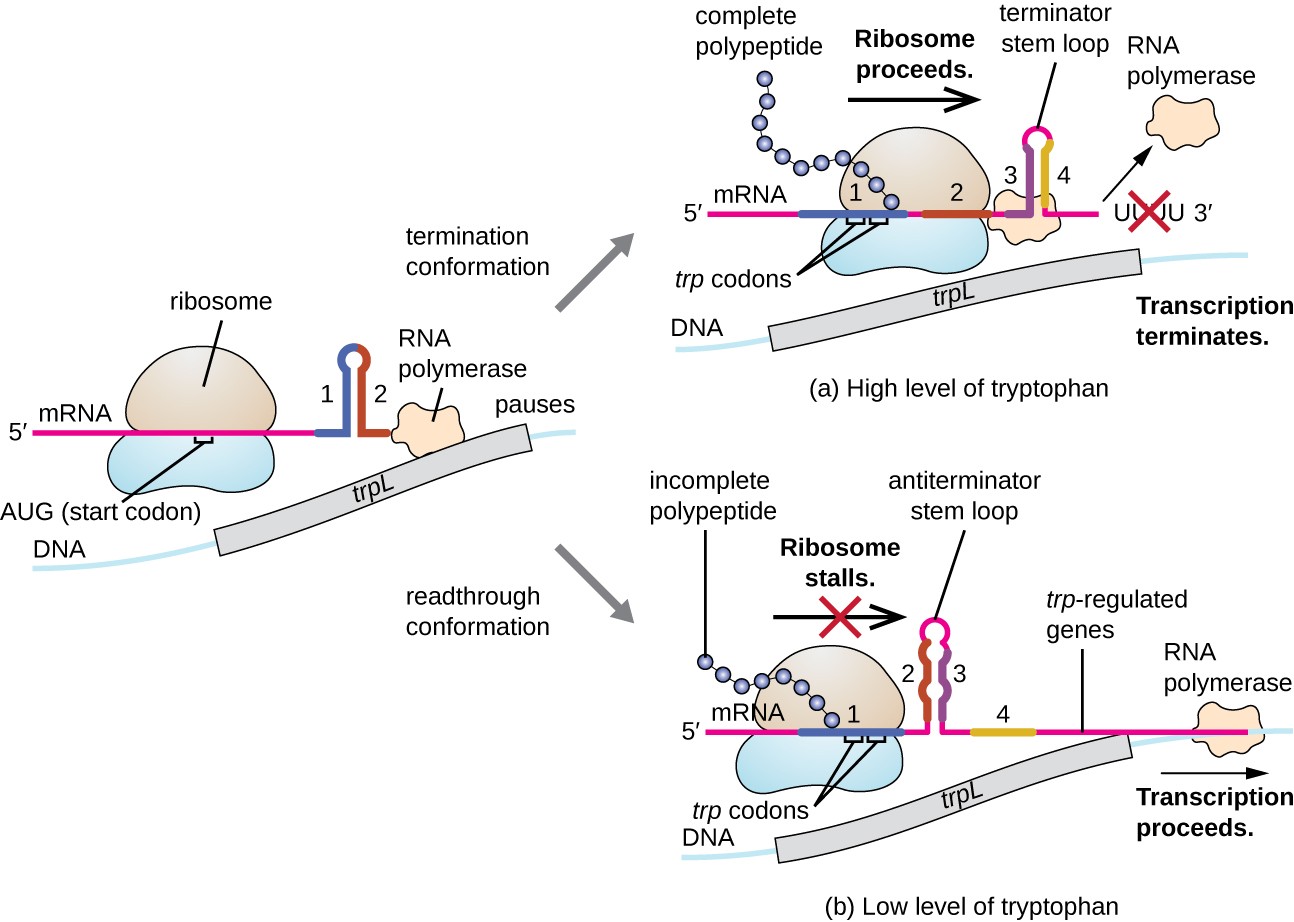
Rysunek 7. Kliknij, aby zobaczyć większy obraz. Gdy tryptofan jest obfity, następuje translacja krótkiego peptydu leader kodowanego przez trpL, powstaje pętla terminatora między regionami 3 i 4, A transkrypcja kończy się., Gdy poziom tryptofanu jest zubożony, translacja krótkiego peptydu leader zatrzymuje się w regionie 1, umożliwiając regionom 2 i 3 tworzenie pętli antyterminatora, a polimeraza RNA może transkrypować geny strukturalne trp operon.
pokrewnym mechanizmem jednoczesnej regulacji transkrypcji i translacji u prokariotów jest użycie ryboswitcha, małego regionu niekodującego RNA znajdującego się w końcu 5′ niektórych cząsteczek prokariotycznego mRNA (ryc. 8). Ryboswitch może wiązać się z małą cząsteczką wewnątrzkomórkową, aby ustabilizować pewne struktury drugorzędowe cząsteczki mRNA., Wiązanie małej cząsteczki determinuje, która struktura pętli macierzystej tworzy, wpływając tym samym na zakończenie syntezy mRNA i syntezy białek.

Rysunek 8. Kliknij, aby powiększyć obraz. Ryboswitche Znalezione w prokariotycznych cząsteczkach mRNA mogą wiązać się z małymi cząsteczkami wewnątrzkomórkowymi, stabilizując pewne struktury RNA, wpływając na zakończenie syntezy samej cząsteczki mRNA (po lewej) lub białka wytwarzanego za pomocą tego mRNA (po prawej).,
inne czynniki wpływające na ekspresję genów w Prokaryotes i Eukaryotes
chociaż skupiamy się na naszej dyskusji nad transcriptional kontroli używane prokaryotic operonów jako przykłady, eukaryotic transcriptional kontroli jest podobny pod wieloma względami. Podobnie jak u prokariotów, transkrypcja eukariotyczna może być kontrolowana poprzez wiązanie czynników transkrypcyjnych, w tym represorów i aktywatorów., Co ciekawe, na transkrypcję eukariotyczną może wpływać wiązanie białek z regionami DNA, zwanymi wzmacniaczami, raczej daleko od genu, poprzez pętlę DNA ułatwioną między wzmacniaczem a promotorem (ryc. 9). Ogólnie rzecz biorąc, regulowanie transkrypcja jest wysoce skutecznym sposobem kontrolować genu wyrażenie w oba prokaryotes i eukaryotes. Jednak, kontrola genu wyrażenie w eukaryotes w odpowiedzi środowiskowy i komórkowy stresy może dokonywać w dodatkowy sposób bez Wiązanie transkrybowanie czynniki region regulacyjny.,

Rysunek 9. U eukariotów enhancer jest sekwencją DNA, która promuje transkrypcję. Każdy wzmacniacz składa się z krótkich sekwencji DNA zwanych dystalnymi elementami kontrolnymi. Aktywatory związane z dystalnymi elementami kontrolnymi oddziałują z białkami mediatorskimi i czynnikami transkrypcyjnymi. Dwa różne geny mogą mieć ten sam promotor, ale różne dystalne elementy kontrolne, umożliwiając różnicową ekspresję genów.,
kontrola poziomu DNA
u eukariotów cząsteczki DNA lub związane z nimi histony mogą być modyfikowane chemicznie w taki sposób, aby wpływać na transkrypcję; nazywa się to regulacją epigenetyczną. Wykazano, że metylacja niektórych nukleotydów cytozyny w DNA w odpowiedzi na czynniki środowiskowe wpływa na wykorzystanie takiego DNA do transkrypcji, przy czym metylacja DNA powszechnie koreluje z obniżonym poziomem ekspresji genów., Dodatkowo, w odpowiedzi na czynniki środowiskowe, histonowe proteiny dla opakowania DNA mogą także chemicznie modyfikować na wiele sposobów, wliczając acetylację i deacetylację, wpływając na pakowanie stan DNA i tym samym wpływając na dostępność luźno rannego DNA dla transkrypcji. Te chemiczne modyfikacje mogą czasami utrzymywać przez wiele rund komórki podział, czyniąc przynajmniej niektóre z tych epigenetycznych zmian dziedzicznych.
Ten film opisuje jak regulacja epigenetyczna kontroluje ekspresję genów.,
zastanów się nad tym
- co zatrzymuje lub pozwala na kontynuowanie transkrypcji, gdy tłumienie działa?
- co decyduje o stanie riboswitcha?
- opisz funkcję wzmacniacza.
- opisują dwa mechanizmy regulacji epigenetycznej u eukariotów.
Clinical Focus: Travis, Resolution
Ten przykład kończy historię Travisa, która rozpoczęła się od funkcji materiału genetycznego, transkrypcji RNA i sposobu, w jaki prokarioty Aseksualne osiągają różnorodność genetyczną.,
chociaż Travis przeżył walkę z martwiczym zapaleniem powięzi, teraz musiałby przejść operację przeszczepu skóry, a następnie długotrwałą fizykoterapię. W oparciu o ilość masy mięśniowej, którą stracił, jest mało prawdopodobne, że jego noga powróci do pełnej siły, ale jego Fizjoterapeuta jest optymistą, że odzyska trochę wykorzystania nogi.
badania laboratoryjne wykazały, że czynnikiem sprawczym zakażenia był szczep paciorkowca z grupy a (ang. „Group a strep”)., Zgodnie z wymogami prawa, sprawa Travisa została zgłoszona do Państwowego Departamentu Zdrowia, a ostatecznie do Centers for Disease Control and Prevention (CDC). W CDC szczep paciorkowca grupy a wyizolowany od Travisa został dokładniej przeanalizowany pod kątem oporności na metycylinę.
oporność na metycylinę jest kodowana genetycznie i staje się coraz bardziej powszechna w paciorkowcach grupy A poprzez horyzontalny transfer genów. W martwiczym zapaleniu powięzi przepływ krwi do zakażonego obszaru jest zazwyczaj ograniczony ze względu na działanie różnych genetycznie zakodowanych toksyn bakteryjnych., Z tego powodu krwawienie w wyniku testu nacięcie jest zazwyczaj niewielkie lub nie występuje. Niestety te toksyny bakteryjne ograniczają skuteczność antybiotyków dożylnych w usuwaniu infekcji ze skóry i tkanek leżących u jej podłoża, co oznacza, że sama oporność na antybiotyki nie tłumaczy nieskuteczności leczenia. Niemniej jednak, antybiotykoterapia dożylna była uzasadniona w celu zminimalizowania możliwego wyniku posocznicy, która jest częstym wynikiem martwiczego zapalenia powięzi., Dzięki analizie genomowej przez CDC szczepu wyizolowanego z Travis wykazano, że kilka ważnych genów wirulencji zostało zakodowanych na prorokach, co wskazuje, że transdukcja jest ważna w poziomym transferze genów tych genów z jednej komórki bakteryjnej do drugiej.
kluczowe pojęcia i podsumowanie
- ekspresja genów jest ściśle regulowanym procesem.
- ekspresja genów u prokariotów jest w dużej mierze regulowana w punkcie transkrypcji. Ekspresja genów u eukariotów jest dodatkowo regulowana po transkrypcji.,
- prokariotyczne geny strukturalne o pokrewnej funkcji są często zorganizowane w operony, wszystkie kontrolowane przez transkrypcję z jednego promotora. Region regulacyjny Operonu obejmuje sam promotor i region otaczający promotor, z którym czynniki transkrypcyjne mogą wiązać się wpływając na transkrypcję.
- chociaż niektóre operony są wyrażane konstytutywnie, większość podlega regulacji poprzez zastosowanie czynników transkrypcyjnych (represorów i aktywatorów)., Represor wiąże się z operatorem, sekwencją DNA w regionie regulacyjnym pomiędzy miejscem wiązania polimerazy RNA w promotorze a pierwszym genem strukturalnym, fizycznie blokując tym samym transkrypcję tych operonów. Aktywator wiąże się w regionie regulacyjnym Operonu, pomagając polimerazie RNA wiązać się z promotorem, zwiększając w ten sposób transkrypcję tego Operonu. Induktor wpływa na transkrypcję poprzez interakcję z tłumikiem lub aktywatorem.
- OPERON trp jest klasycznym przykładem Operonu represyjnego., Gdy tryptofan gromadzi się, tryptofan wiąże się z represorem, który następnie wiąże się z operatorem, uniemożliwiając dalszą transkrypcję.
- lac operon jest klasycznym przykładem indukcyjnego Operonu. Gdy laktoza jest obecna w komórce, jest przekształcana w allolaktozę. Allolaktoza działa jako induktor, wiążąc się z represorem i uniemożliwiając represorowi Wiązanie się z operatorem. Pozwala to na transkrypcję genów strukturalnych.
- lac operon również podlega aktywacji., Gdy poziom glukozy jest zubożony, część komórkowego ATP jest przekształcana w cAMP, który wiąże się z białkiem aktywatora katabolitu (CAP). Kompleks Camp-CAP aktywuje transkrypcję lac operon. Gdy poziom glukozy jest wysoki, jego obecność uniemożliwia transkrypcję lac Operonu i innych operonów przez katabolizm.
- małe cząsteczki wewnątrzkomórkowe zwane alarmonami powstają w odpowiedzi na różne naprężenia środowiskowe, umożliwiając bakteriom kontrolowanie transkrypcji grupy operonów, zwanych regulonami.,
- bakterie mają zdolność do zmiany, którego σ czynnika polimerazy RNA używają w odpowiedzi na warunki środowiskowe, aby szybko i globalnie zmienić, które regulony są transkrybowane.
- Prokaryoty mają mechanizmy regulacyjne, w tym tłumienie i stosowanie ryboswitchów, aby jednocześnie kontrolować zakończenie transkrypcji i tłumaczenie z tego transkryptu. Mechanizmy te działają poprzez tworzenie pętli macierzystych w końcu 5′ cząsteczki mRNA obecnie syntetyzowanej.,
- istnieją dodatkowe punkty regulacji ekspresji genów u prokariotów i eukariotów. U eukariotów Regulacja epigenetyczna poprzez chemiczną modyfikację DNA lub histonów oraz regulacja przetwarzania RNA to dwie metody.
wielokrotnego wyboru
operon genów kodujących enzymy w szlaku biosyntetycznym może być który z poniższych?
- inducible
- repressible
- constitutive
- monocistronic
o genach kodujących operon, które są transkrybowane i tłumaczone w sposób ciągły, aby zapewnić komórce stały pośredni poziom produktów białkowych mówi się, który z poniższych?
- repressible
- inducible
- constitutive
- activated
który z poniższych warunków prowadzi do maksymalnej ekspresji lac operon?
- laktoza obecna, glukoza nieobecna
- laktoza obecna, glukoza nieobecna
- laktoza nieobecna, glukoza nieobecna
- laktoza nieobecna, glukoza nieobecna
który z poniższych typów regulacji ekspresji genów jest unikalny dla eukariotów?,
- tłumienie
- wykorzystanie alternatywnego czynnika σ
- chemiczna modyfikacja histonów
- alarmony
wypełnij puste pole
sekwencja DNA, z którą mogą wiązać się represory, leżąca pomiędzy promotorem a pierwszym genem strukturalnym, nazywa się ________.,
zapobieganie ekspresji operonów kodujących podłoże dla substratów innych niż glukoza w przypadku obecności glukozy nazywa się_______.
pomyśl o tym
- jakie są dwa sposoby, w jakie bakterie mogą wpływać na transkrypcję wielu różnych operonów jednocześnie w odpowiedzi na określony stan środowiska?
- poniższy rysunek pochodzi z oryginalnej pracy Monod dotyczącej wzrostu diauxycznego, pokazującej wzrost E. coli w jednoczesnej obecności ksylozy i glukozy jako jedynego źródła węgla. Wyjaśnij, co dzieje się w punktach A–D w odniesieniu do źródła węgla używanego do wzrostu i wyjaśnij, czy wyrażany jest operon wykorzystujący ksylozę (i dlaczego)., Należy zauważyć, że ekspresja enzymów wymaganych do stosowania ksylozy jest regulowana w sposób podobny do ekspresji enzymów wymaganych do stosowania laktozy.
