Eukaryotické Struktury Chromozomů
délka DNA v jádře je daleko větší, než je velikost compartmentin kterém je obsažen. Aby se vešla do tohoto oddílu, musí být DNA nějakým způsobem uvolněna. Stupeň, do kterého je dna kondenzována, je vyjádřenjako jeho obalový poměr.
Balení poměru – délka DNA rozděleny podle délky, do které je zabalen
například, nejkratší lidský chromozom obsahuje 4,6 x 107 bp DNA(asi 10 krát velikost genomu E. coli)., To odpovídá 14 000 µm rozšířené DNA. Ve svém nejvíce kondenzovaném stavu během mitózychromozom je asi 2 µm dlouhý. To dává poměr balení 7000 (14 000/2).
pro dosažení celkového balicího poměru není DNA balena přímo do konečné struktury chromatinu. Místo toho obsahuje několik hierarchií organizace.První úroveň balení je dosaženo tím, že vinutí DNA kolem proteincore produkovat „korálek-like“ struktury, zvané nucleosome. To je poměr balení asi 6., Tato struktura je invariantní v obouteuchromatin a heterochromatin všech chromozomů. Druhá úroveň baleníje stáčení korálků v spirálové struktuře nazývané Fiber 30 nm, který se nachází jak v chromatinu interfázy, tak v mitotických chromozomech. Tím se zvyšuje poměr balení na cca 40. Konečné balení occurswhen vlákno je organizováno do smyček, lešení a domén, které dávají finalpacking poměr asi 1000 v interfázových chromozomů a asi 10.000 inmitotic chromozomů.,
eukaryotické chromozomy se skládají z komplexu DNA-bílkovin, který je organizován kompaktním způsobem, který umožňuje uložení velkého množství DNA v jádru buňky. Podjednotkové označení chromozomu je chromatin.Základní jednotkou chromatinu je nukleosom.,romatin – jednotka analýzy chromozomu; chromatin reflectsthe obecné struktury chromozomu, ale nejsou jedinečné na jakékoliv particularchromosome
Nucleosome – nejjednodušší obaly, struktury DNA, která se nachází ve všech eukaryotických chromozomů; DNA je zabalena kolem octamer malých basicproteins zvané histony; 146 bp je omotal kolem jádra a remainingbases odkaz na další nucleosome; tato struktura způsobuje negativní supercoiling
nucleosome se skládá z asi 200 bp omotané kolem histonů octamerthat obsahuje dvě kopie proteinů histonů H2A, H2B, H3 a H4., Ty jsou známé jako základní histony. Histony jsou základní proteiny, které mají afinitupro DNA a jsou nejhojnějšími bílkovinami spojenými s DNA. Aminoacidní sekvence těchto čtyř histonů je konzervována, což naznačuje podobnou funkcipro všechny.
Délka DNA, která je spojena s nukleosomovou jednotkou, se liší mezidruh. Ale bez ohledu na velikost se jedná o dvě složky DNA. CoreDNA je DNA, která je ve skutečnosti spojena s histonovým oktamerem.Tato hodnota je invariantní a je 146 párů bází., Jádro DNA tvoří dvě smyčkykolem oktameru, a to umožňuje dvě oblasti, které jsou 80 BP od sebe tobe přivedl do těsné blízkosti. Takže dvě sekvence, které jsou daleko od sebe, mohouinterakt se stejným regulačním proteinem pro kontrolu genové exprese. Dna, která je mezi každou histonů octamer je tzv. linker DNAand se mohou lišit v délce od 8 do 114 párů bází. Tato variace je druhapecifická, ale změna délky linker DNA byla také spojena s vývojovým stupněm organismu nebo specifickými oblastmi genomu.,
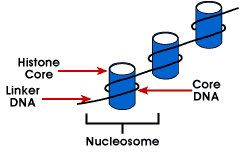
další úroveň organizace chromatinu je 30 nm vlákno. Jedná se o solenoidovou strukturu s asi 6 nukleosomy na otáčku. To je poměr balení 40, což znamená, že každých 1 µm podél axisobsahuje 40 µm DNA. Stabilita této struktury vyžadujepřítomnost posledního člena rodiny genů histonu, histone H1., Becauseexperiments, že pás H1 z chromatinu udržení nucleosome, ale ne 30 nm strukturu, se dospělo k závěru, že H1 je důležité pro stabilizationof 30 nm strukturu.
konečná úroveň balení je charakterizována strukturou 700 nm viděnou v chromozomu metafázy. Kondenzovaný kus chromatinu má acharakteristickou strukturu lešení, kterou lze detekovat v metafasechromosomech. Zdá se, že je to výsledek rozsáhlého smyčkování dnna chromozomu.
poslední definice, které je třeba uvést, jsou euchromatin aheterochromatin., Když jsou chromozomy obarveny barvivy, zdá se, že majístřídavé lehce a tmavě zbarvené oblasti. Lehce zbarvené regionyjsou euchromatin a obsahují jednokopiovou geneticky aktivní DNA. Thetmavě zbarvené oblasti jsou heterochromatin a obsahují opakující se sekvencekteré jsou geneticky neaktivní.
centromery a telomery
centromery a telomery jsou dvěma základními rysy všech eukaryotických chromozomů. Každá z nich poskytuje jedinečnou funkci, která je naprosto nezbytná pro stabilitu chromozomu., Centromery jsou vyžadovány pro segregaci centromeru během meiózy a mitózy a teleomery poskytují terminální stabilitu chromozomu a zajišťují jeho přežití.
centromery jsou ty kondenzované oblasti uvnitř chromozomu, které jsou zodpovědné za přesnou segregaci replikované chromozomedurující mitózy a meiózy. Když jsou chromozomy obarveny, obvykle vykazují tmavě zbarvenou oblast, která je centromerem. Během mitózy se centromere, který sdílí sesterské chromatidy, musí rozdělit tak, aby chromatidy mohly migrovatopačné póly buňky., Na druhou stranu, během prvního meiotického rozdělenícentromer sesterských chromatidů musí zůstat neporušený, zatímco během meiosisII musí jednat stejně jako během mitózy. Centromere je tedy důležitou složkou chromozomové struktury a segregace.
V centromera oblast, většina druhů má několik míst, kde vřeteno vlákna připojit, a tyto stránky se skládají z DNA, stejně jako protein. Skutečné místo, kde dochází k připojení, se nazývá kinetochore a skládá se z DNA i bílkovin. Sekvence DNA v těchto oblastech je nazývánacen DNA., Protože cen DNA může být přesunuta z jednoho chromozomu do druhého a stále poskytuje chromozomu schopnost segregovat, tyto sekvence nesmí poskytovat žádnou jinou funkci.
Typicky CEN DNA je o 120 párů bází dlouhý a skládá se z několika dílčích sub-domén, CDE-I, CDE-II a CDE-III. Mutace v prvních dvou sub-domén, nemají žádný vliv na segregaci, ale o bodovou mutaci v CDE-III sub-domény, zcela eliminuje schopnost centromerou funkce při segregaci chromozomů., Proto se CDE-III musí aktivně podílet na vazbě vřetenových vláken na centromere.
proteinová složka kinetochoru se nyní vyznačuje. Komplex tří proteinů zvaných Cbf-III se váže na normální oblasti CDE-III, ale nemůže se vázat na oblast CDE-III s bodovou mutací, která zabraňuje mitotické segregaci. Kromě toho mutanty genů kódujících proteiny Cbf-III také eliminují schopnost chromozomů segregovat se během mitózy., Další analýzy DNA a proteinových složek centromere jsou nezbytné k úplnému pochopení mechaniky segregace chromozomů.
telomery jsou oblast DNA na konci lineárního eukaryotického chromosomu, které jsou potřebné pro replikaci a stabilitu chromozomu.McClintock uznal jejich zvláštní vlastnosti, když si všimla, že pokud twochromosomes byly rozděleny v buňce, na konci jeden mohl připojit k zdůraznila, naopak. To, co nikdy nepozorovala, bylo připevnění zlomeného ke konci neporušeného chromozomu., Tak konce zlomených chromozomůjsou lepkavé, zatímco normální konec není lepkavý, což naznačuje, že koncechromosomy mají jedinečné vlastnosti. Obvykle, ale ne vždy, telomeridna je heterochromatická a obsahuje přímé tandemly opakované sekvence. Následující tabulka ukazuje opakované sekvence několika druhů. Jedná se často o formu (T / a) xGy, kde x je mezi 1 a 4 a y je větší než 1.,
Telomere Repeat Sequences
| Species | Repeat Sequence |
|---|---|
| Arabidopsis | TTTAGGG |
| Human | TTAGGG |
| Oxytricha | TTTTGGGG |
| Slime Mold | TAGGG |
| Tetrahymena | TTGGGG |
| Trypanosome | TAGGG |
| Yeast | (TG)1-3TG2-3 |
Notice that the number of TG sequences and the number of cytosines in the yeast sequence varies., Alespoň u kvasinek bylo prokázáno, že různé kmeny obsahují různé délky teleomerů a že délka je pod genetickou kontrolou.
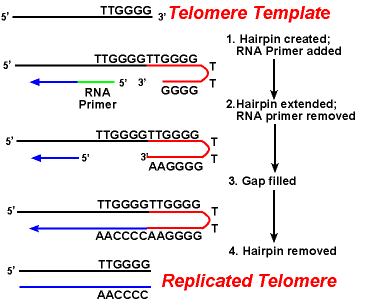
primárním problémem telomer je replikace zaostávajícího pramene.Protože syntéza DNA vyžaduje RNA šablony (která poskytuje volný 3′-oh skupinu) na prime replikace DNA, a tato šablona je nakonec degradován, krátké jednořetězcové oblasti by odešel na konci chromozomu. Tato oblast by byla náchylná k enzymům, které degradují jednovláknovou DNA., Výsledkem by bylo, že délka chromozomu by se po každém rozdělení zkrátila. Ale to není vidět.
působení enzymů telomerázy zajišťuje správnou replikaci konců zaostávajících pramenů. Dobře studovaný systém zahrnuje organismus Tetrahymena protozoa. Telomery tohoto organismu končí v sekvenci 5 ‚- TTGGGG-3‘. Telomeráza přidává sérii opakování 5′-TTGGG-3 ‚ na konce zaostávajícího pramene. Vlásenka nastane, když neobvyklé páry bází mezi zbytky guaninu v opakované formě., Dále je odstraněn primer RNA a 5′ konec zaostávajícího pramene může být použit pro syntézu DNA. Ligace nastává mezi hotovým zaostávajícím pramenem a vlásenkou. Nakonec se vlásenka odstraní při opakování 5′-TTGGG-3‘. Tak je konec chromozomu věrně replikován. Následující obrázek ukazuje tyto kroky.,
Replikace Telomer
Analýza Sekvencí DNA v Eukaryotických Genomů
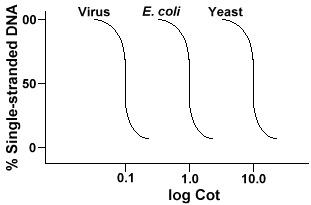
techniku, která se používá k určení sekvence složitost genomeinvolves denaturace a renaturation DNA. DNA je denaturována zahříváním, které roztaví H-vazby a vykreslí dna jednovláknovou. Pokud se DNA rychle ochladí, DNA zůstává jednovláknová. Pokud se však DNAis nechá pomalu ochladit,sekvence, které se doplňují, se najdou a nakonec se znovu spojí., Rychlost, s jakou DNA reanneals (jiný termín pro renature) je funkce druhu, ze kterého DNAwas izolované. Níže je křivka, která se získává z jednoduchého genomu.
osa Y je procento DNA, která zůstává jednovláknová. To je vyjádřeno jako poměr koncentrace jednovláknové DNA (C) k celkové koncentraci počáteční DNA (Co). Osa X je log-měřítko produktu počáteční koncentrace DNA (v molech/litrech)vynásobené délkou času reakce pokračovala (v sekundách)., Označení pro tuto hodnotu je dětská postýlka a nazývá se hodnota „Dětská postýlka“. Samotná křivka se nazývá křivka „postýlky“. Jak je vidět, křivka je poměrně hladká, což naznačuje, že k opětovnému žíhání dochází zpomaleně, ale postupně po určitou dobu. Jedna konkrétní hodnota, která je užitečná, je Cot½, hodnota cot, kde se znovu objevila polovina DNA.
kroky spojené s denaturací DNA a Renaturací
1. Zkraťte DNA na velikost asi 400 bp.
2. Denaturujte DNA zahřátím na 100oC.
3. Pomalu ochlaďte a odeberte vzorky v různých časových intervalech.
4., Určete % jednovláknovou DNA v každém časovém bodě.
tvaru „Postýlky“ křivky pro daný druh, je funkce dvou faktorech:
- velikost nebo složitost genomu; a
- množství repetitivní DNA v genomu
Když jsme plot „Postýlka“ křivky genomu ze tří druhů, jako jsou bacteriophagelambda, E. coli a kvasinek budeme vidět, že mají stejný tvar,ale Cot½ kvasinek bude největší, E. coli nextand lambda nejmenší., Fyzicky, větší velikost genomu tím déle bude trvat pro každou sekvenci se setkáte s jeho komplementární sekvence v řešení. Je to proto, že se musí setkat dvě komplementární sekvencekaždý jiný, než se mohou spárovat. Čím složitější je genom, tím více unikátních sekvencí, které jsou k dispozici, tím déle bude trvat, než se anydvě komplementární sekvence setkají a spárují. Vzhledem k podobným situacím v roztoku pak bude trvat složitější druh, který bude mít delší dosah .,
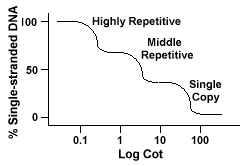
Opakované DNA sekvence, sekvence DNA, které se vyskytují více než jednou v genomu druhu, mají výrazné účinky na „Postýlku“ křivky.Pokud konkrétní sekvence je zastoupena dvakrát v genomu bude mít twocomplementary sekvence spárovat s a jako takový bude mít Postýlku hodnota halfas velké jako posloupnost zastoupena pouze jednou v genomu.
eukaryotické genomy mají ve skutečnosti širokou škálu sekvencí, které jsou reprezentovány na různých úrovních opakování., Sekvence jedné kopie se nacházejí jednounebo několikrát v genomu. Mnoho sekvencí, které kódují funkcialgenes spadají do této třídy. Střední opakující se DNA se nacházejí od 10s -1000 krát v genomu. Příklady z nich by zahrnovaly rRNA a trnageny a skladovací proteiny v rostlinách, jako je kukuřice. Střední opakující se DNAcan se pohybuje od 100-300 bp do 5000 bp a může být rozptýlen v celém genomu.Nejhojnější sekvence se nacházejí ve vysoce opakujících se DNAclass., Tyto sekvence se nacházejí od 100 000 do 1 milionkrát v genomea může se pohybovat ve velikosti od několika do několika set bází na délku. Tyto jevy se vyskytují v oblastech chromozomu, jako je heterochromatin, centromery a telomery, a mají tendenci být uspořádány jako tandemové opakování. Následující je příklad tandemly opakované sekvence:
ATTATA ATTATA ATTATA // ATTATA
Genomů, které obsahují tyto různé třídy sekvence reanneal v differentmanner než genomy se pouze jedna kopie sekvence., Místo toho, jeden hladký „Postýlka“ křivka, tři odlišné křivky lze vidět, každý představuje jiný opakování třídy. První sekvence, které se znovu objeví, jsou vysokérepetitivní sekvence, protože v genomu existuje tolik kopií a protože mají nízkou sekvenční složitost. Druhá část genomu k reanneal je střední opakující se DNA a poslední část toreanneal je jediná kopie DNA., Následující diagram zobrazuje „Postýlka“křivka pro „typické“ eukaryotický genom,
V následující tabulce jsou uvedeny sekvence distribuce pro vybrané druhy.
| druh | |
|---|---|
| bakterie | 99.,27% Střední Opakující se 8% Vysoce Opakující se |
| Kukuřice | 30% Jediném exempláři 40% Střední Opakující se 20% Vysoce Opakující se |
| Pšenice | 10% Jediném exempláři 83% Střední Opakující se 4% Vysoce Opakující se |
| Arabidopsis | 55% Jediném exempláři 27% Střední Opakující se 10% Vysoce Opakující se |
Pořadí Interspersion
I když genomech vyšších organismů obsahují jednu kopii, middlerepetitive a vysoce repetitivní sekvence DNA, tyto sekvence nejsou arrangedsimilarly ve všech druhů., Prominentní uspořádání se nazývá krátké obdobíintersperze. Toto uspořádání je charakterizováno opakovanými sekvencemi100 – 200 bp v délce rozptýlené mezi jednotlivými kopírovacími sekvencemi, které mají délku 1000-2000bp. Toto uspořádání se vyskytuje u zvířat, hub a rostlin.
druhým typem uspořádání je dlouhodobá intersperze. To je charakterizováno 5000 BP úseky opakovaných sekvencí protínaných v oblastech 35 000 bp jednokopiové DNA. Drosophila je příkladdruh s tímto neobvyklým uspořádáním sekvence., V obou případech jsou sekvence obvykle ze střední opakující se třídy. Diskutovali jsme o tom, kde se nacházejí vysoce opakující se sekvence.
Eukaryotický chromozom karyotyp
zatímco bakterie mají pouze jeden chromozom, eukaryotické druhy mají alespoň jeden pár chromozomů. Většina z nich má více než jeden pár. Dalším relevantním bodem je, že eukaryotické chromozomy jsou detekovány pouze během celldivision a ne během všech fází buněčného cyklu. Jsou ve své nejvícekondenzované formě během metafázy, když jsou připojeny sesterské chromatidy.,Toto je primární fáze, kdy se provádí cytogenetická analýza.
každý druh je charakterizován karyotypem. Karyotyp je adescripce počtu chromozomů v normální diploidní buňce, stejně jakojako jejich rozdělení velikosti. Například lidský chromozom má 23 párů chromozomu, 22 somatických párů a jeden pár pohlavních chromozomů. Jedním z důležitýchaspekt genetického výzkumu je korelace změn karyotypu se změnamiv fenotypu jednotlivce.
jedním z důležitých aspektů genetiky je korelace změn karyotypuzměny ve fenotypu., Například lidé, kteří mají extra chromozom 21mají Downův syndrom. Inzerce, delece a změny v počtu chromozomůmůže být detekován kvalifikovaným cytogenetikem, ale jejich korelace specifické fenotypy jsou obtížné.
prvním diskriminačním parametrem při vývoji karyotypu je velikosta počet chromozomů. I když je to užitečné, neposkytujedostatek detailů, které mají být zahájeny vývoj korelace mezi strukturaa funkce (fenotyp)., Chcete-li dále rozlišovat mezi chromozomy, onijsou ošetřeny barvivem, které reprodukuje DNA reprodukovatelným způsobem. Po barvení jsou některé regiony lehce obarvené a jiné jsou těžké. Jak bylo popsáno výše, lehce obarví regiony jsou calledeuchromatin, a tmavě zbarvená oblast je calledheterochromatin. Současným barvivem je skvrna Giemsa a výsledný vzor se nazývá G-banding vzor.,
C-value Paradox
kromě popisu genomu organismu počtem chromozomů je také popsán množstvím DNA v haploidní buňce. To je usuallyexpressed jako množství DNA na haploidní buňky (obvykle vyjádřené jako pikogramů)nebo počet kilobází na haploidní buňky a nazývá se C hodnotu. Okamžitým funkce eukaryotických organismů upozorňuje na zvláštní anomálii, bylo včasné odhalení v molekulární výzkum., I když eukaryotické organismyzačnou mít 2-10krát více genů než prokaryoty, mají v buňce mnoho řádůmnožství více DNA. Navíc množství DNA na genomje korelována s předpokládanou evoluční složitostí druhu.To je uvedeno jako paradox hodnoty C: množství DNA v haploidcellu organismu nesouvisí s jeho evoluční složitostí. (Dalším důležitým bodem, který je třeba mít na paměti, je, že mezi počtem chromozomů a předpokládanou evoluční složitostí anorganismu neexistuje žádný vztah.,)
C Values of Organisms Used in Genetic Studies
| Species | Kilobases/haploid genome |
|---|---|
| E. coli |
4.5 x 103 |
| Human |
3.0 x 106 |
| Drosophila |
1.7 x 105 |
| Maize |
2.0 x 106 |
| Aribidopsis |
7.,0 x 104 |
dramatický příklad rozsahu C hodnoty lze vidět v království, kde výrobní závod Arabidopsis představuje low-end a lily (1, 0 x 10^8 kb/haploidgenome) high-end složitost. Z hlediska hmotnosti je to 0,07 picogramsper haploidní Arabidopsis genom a 100 pikogramů na haploidní lilygenom.
genom-kompletní sada chromozomů zděděných od jednoho rodiče; kompletní složka dna jednotlivce; definice často vylučujeorganely