eukarióta kromoszóma szerkezete
a DNS hossza a magban sokkal nagyobb, mint a benne lévő kompartment mérete. Ahhoz, hogy beilleszkedjen ebbe a rekeszbe, a DNS-nek kell lenniebizonyos módon érzékelik. A DNS kondenzációjának mértéke kifejeződikmint a csomagolási aránya.
csomagolási arány-a DNS hossza osztva a hosszávalcsomagolva
például a legrövidebb emberi kromoszóma 4,6 x 107 bp DNS-t tartalmaz (az E. coli genomméretének körülbelül 10-szerese)., Ez 14 000 µm kiterjesztett DNS-nek felel meg. A mitózis során a leginkább kondenzált állapotbana kromoszóma körülbelül 2 µm hosszú. Ez 7000(14 000/2) csomagolási arányt biztosít.
a teljes csomagolási arány elérése érdekében a DNS-t nem csomagolják közvetlenül a kromatin végleges szerkezetébe. Ehelyett több szervezeti hierarchiát tartalmaz.A csomagolás első szintjét úgy érik el, hogy a DNS-t egy protein körül tekercselik, hogy egy “gyöngyszerű” szerkezetet hozzanak létre, amelyet nukleoszómának neveznek. Ez körülbelül 6 csomagolási arányt biztosít., Ez a szerkezet invariáns mind aaz összes kromoszóma euchromatinja és heterokromatinja. A csomagolás második szintjea gyöngyök tekercselése egy spirális szerkezetben, a 30 nm-es fiberben, amely mind az interfáz-kromatinban, mind a mitotikus kromoszómákban megtalálható. Eza szerkezet körülbelül 40-re növeli a csomagolási arányt. A végső csomagolás akkor következik be, amikor a szál hurkokba, állványokba és doménekbe van rendezve, amelyek az interphase kromoszómákban kb. 1000, az inmitotikus kromoszómákban pedig kb. 10 000 inmitotikus kromoszómát adnak.,
Az eukarióta kromoszómák egy olyan DNS-fehérje komplexből állnak, amely kompakt módon szerveződik, amely lehetővé teszi a nagy mennyiségű DNS tárolását a sejtmagban. A kromoszóma alegysége a kromatin.A kromatin alapvető egysége a nukleoszóma.,romatin – az elemzés egysége a kromoszóma; kromatin reflectsthe általános szerkezete a kromoszóma de nem egyedülálló particularchromosome
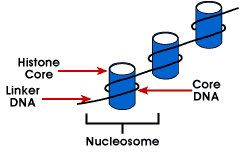
Nucleosome – legegyszerűbb csomagolás szerkezete DNS-t, amely megtalálható minden eukarióta kromoszómák; DNS köré egy octamer kis basicproteins úgynevezett histones; 146 bp köré a core, valamint a remainingbases link a következő nucleosome; ez a struktúra okozza a negatív supercoiling
A nucleosome áll, körülbelül 200 bp köré egy histone octamerthat tartalmaz két példányban histone fehérjék H2A, H2B, H3, valamint H4., Ezek a fő hisztonok. A hisztonok olyan alapvető fehérjék, amelyeknek affinitása van a DNS-hez, és a DNS-hez kapcsolódó leggyakoribb fehérjék. Ennek a négy hisztonnak az aminoacid szekvenciája megmarad, ami mindenki számára hasonló funkciót javasol.
A DNS hossza, amely a nukleoszómaegységhez kapcsolódik, a fajok között változik. De a mérettől függetlenül két DNS-összetevő vesz részt. A CoreDNA az a DNS, amely valójában a hiszton-oktamerhez kapcsolódik.Ez az érték invariáns és 146 bázispár., A mag DNS-e két ciklust alkot az oktamer körül, és ez lehetővé teszi, hogy két, egymástól 80 bp-re eső régió közel kerüljön egymáshoz. Így két egymástól távol lévő szekvencia képesugyanazzal a szabályozó fehérjével kölcsönhatásba léphet a génexpresszió szabályozására. TheDNA, amely az egyes hiszton oktamer nevezzük a linker DNAand hossza változhat 8-114 bázis Párok. Ez a variáció fajspecifikus, de a linker DNS hosszának változása is összefügga szervezet fejlődési stádiuma vagy a genom specifikus régiói.,
a kromatin szervezésének következő szintje a 30 nm-es szál. Ezfordulatonként körülbelül 6 nukleoszómával rendelkező mágnesszelep-szerkezetnek tűnik. Ez 40 csomagolási arányt biztosít, ami azt jelenti, hogy az Axis mentén minden 1 µm 40 µm DNS-t tartalmaz. Ennek a szerkezetnek a stabilitása megkövetelia hiszton géncsalád utolsó tagjának, a hiszton H1-nek a jelenléte., Mivel a H1-et kromatinból leválasztó kísérletek fenntartják a nukleoszómát, de nema 30 nm-es szerkezet, arra a következtetésre jutottak, hogy a H1 fontos a 30 nm-es szerkezet stabilizálásához.
a csomagolás végső szintjét a metafázis kromoszómában látható 700 nm-es szerkezet jellemzi. A kromatin kondenzált darabja akarakterisztikus állványszerkezet, amely metafaszekromoszómákban kimutatható. Úgy tűnik, hogy ez a dnain kromoszóma kiterjedt hurkolásának eredménye.
az utolsó meghatározások, amelyeket be kell mutatni, az euchromatin ésheterokromatin., Amikor a kromoszómákat színezékekkel festik, úgy tűnik, hogy enyhén és sötéten foltos régiók vannak. Az enyhén foltos területek euchromatin, és egypéldányos, genetikailag aktív DNS-t tartalmaznak. A sötét foltos területek heterokromatinok, ismétlődő szekvenciákat tartalmaznak, amelyek genetikailag inaktívak.
Centromeres és Telomeres
Centromeres és telomeres az összes eukarióta kromoszóma két alapvető jellemzője. Mindegyik egyedülálló funkciót biztosít, amely feltétlenül szükséges a kromoszóma stabilitásához., A centromerek a meiózis és mitózis során a centromerek elkülönítéséhez szükségesek, a teleomerek pedig a kromoszóma terminális stabilitását biztosítják, és biztosítják annak túlélését.
a centromerek azok a kondenzált régiók a kromoszómán belül, amelyek felelősek a replikált kromoszóma pontos elkülönítéséért a mitózis és a meiózis során. Amikor a kromoszómákat festik, általában sötét foltos régiót mutatnak, amely a centromere. A mitózis során a testvérkromatidok által megosztott centromernek meg kell osztódnia, hogy a kromatidok átkerülhessenek a sejtpozit pólusai közé., Másrészt az első meiotikus Osztás sorána nővérkromatidok centromere sértetlennek kell maradnia, míg a meiosisII alatt ugyanúgy kell működniük, mint a mitózis során. Ezért a centromere a kromoszómaszerkezet és a szegregáció fontos eleme.
a centromere régióban a legtöbb fajnak több helye van, ahol az orsószálak kapcsolódnak, és ezek a helyek DNS-ből, valamint fehérjéből állnak. A kötődés tényleges helyét kinetochore-nak nevezik, amely mind DNS-ből, mind fehérjéből áll. Ezekben a régiókban a DNS-szekvenciát hívjákcen DNS., Mivel a CEN DNS áthelyezhető egyik kromoszómáról a másikra, és továbbra is biztosítja a kromoszómának a szétválasztás képességét, ezek a szekvenciák nem nyújthatnak más funkciót.
Jellemzően CEN DNS mintegy 120 bázispár hosszú áll, több al-domain, CDE-én, CDE-II. CDE-III. Mutációk az első két al-domain nincs hatással a szegregáció, de egy pontmutáció a CDE-III-al-domain teljesen kiküszöböli a képesség, a centromere működése során kromoszóma szegregáció., Ezért a CDE-III-nak aktívan részt kell vennie az orsószálak centromérhoz való kötésében.
a kinetochore fehérjeösszetevőjét csak most jellemzik. A Cbf-III nevű három fehérje komplexe kötődik a normál CDE-III régiókhoz, de nem képes kötődni egy CDE-III régióhoz olyan pontmutációval, amely megakadályozza a mitotikus szegregációt. Ezenkívül a Cbf-III fehérjéket kódoló gének mutánsai kiküszöbölik a kromoszómák szétválasztásának képességét a mitózis során., A centromere DNS-és fehérjeösszetevőinek további elemzésére van szükség a kromoszóma szegregáció mechanikájának teljes megértéséhez.
a telomerek a DNS régiója a kromoszóma replikációjához és stabilitásához szükséges lineáris eukarióta kromoszóma végén.McClintock felismerte sajátosságait, amikor észrevette, hogy ha kétkromoszómák törnek egy cellában, az egyik vége a másikhoz kapcsolódhatés fordítva. Amit soha nem figyelt meg, az a brokenendnek a megszakítatlan kromoszóma végéhez való kötődése volt., Így a törött kromoszómák végei ragadósak, míg a normál vég nem ragadós, ami arra utal, hogy a kromoszómák végei egyedi tulajdonságokkal rendelkeznek. Általában, de nem mindig, a telomeridna heterokromatikus, közvetlen tandemly ismétlődő szekvenciákat tartalmaz. A következő táblázat több faj ismétlődő sorozatát mutatja. Ezek gyakran az xgy (T/A)formájúak, ahol x 1 és 4 között van, y pedig 1-nél nagyobb.,
Telomere Repeat Sequences
| Species | Repeat Sequence |
|---|---|
| Arabidopsis | TTTAGGG |
| Human | TTAGGG |
| Oxytricha | TTTTGGGG |
| Slime Mold | TAGGG |
| Tetrahymena | TTGGGG |
| Trypanosome | TAGGG |
| Yeast | (TG)1-3TG2-3 |
Notice that the number of TG sequences and the number of cytosines in the yeast sequence varies., Legalább az élesztő esetében kimutatták, hogy a különböző törzsek különböző hosszúságú teleomereket tartalmaznak, és hogy a hossza genetikai ellenőrzés alatt áll.
a telomerek elsődleges nehézsége a lemaradó szál replikációja.Mivel a DNS-szintézis RNS-sablont igényel (amely a szabad 3 ‘ -OHgroup-ot biztosítja) a DNS-replikáció príméhez, és ez a sablon végül lebomlik, egy rövid, egyszálú régió maradna a kromoszóma végén. Ez a régió érzékeny lenne az enzimekre, amelyek lebontják az egyszálú DNS-t., Az eredmény az lenne, hogy a kromoszóma hossza minden osztás után lerövidül. De ez nem látható.
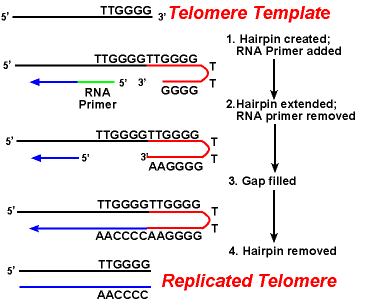
a telomeráz enzimek hatása biztosítja, hogy a lemaradó szálak végei helyesen replikálódjanak. Egy jól tanulmányozott rendszer magában foglalja a Tetrahymena protozoa szervezetet. Ennek a szervezetnek a telomerei az 5′-TTGGGGG-3 ‘ szekvenciában végződnek. A telomeráz egy sor 5′-TTGGGG-3’ ismétlést ad a lemaradó szál végéhez. A hajtű akkor fordul elő, amikor a guaninmaradványok között szokatlan bázispár van az ismétlődő formában., Ezután eltávolítják az RNS-alapozót, majd a lemaradó szál 5 ‘ vége felhasználható DNS-szintézisre. Ligálás történik a kész lag szál és a hajtű között. Végül a hajtűt eltávolítjuk az 5 ‘- TTGGGG-3 ‘ ismétlésnél. Így a kromoszóma vége hűen replikálódik. Az alábbi ábra ezeket a lépéseket mutatja.,
a telomerek replikációja
Az eukarióta genomokban lévő DNS-szekvenciák elemzése
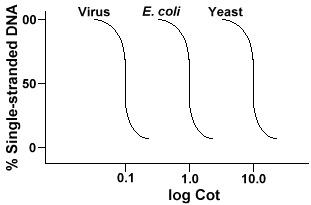
bármely genomeinv szekvencia összetettségének meghatározására használt technika a DNS denaturálása és renaturálása. A DNS-t denatured hevítéssel olvasztják, ami megolvasztja a H-kötéseket, és a DNS-t egyszálúvá teszi. Haa DNS gyorsan lehűl, a DNS egyszálú marad. De ha a DNAis hagyjuk lehűlni lassan, szekvenciák, amelyek kiegészítik fogja találni egymást, majd végül bázis pár újra., Az arány, amellyel a DNS reanneals(egy másik kifejezés renature) egy függvény a faj, amelyből a DNAwas izolált. Az alábbiakban egy egyszerű genomból nyert görbe található.
Az Y-tengely az egyetlen szálú DNS százaléka. Ez az egyszálú DNS (C)koncentrációjának a kiindulási DNS (Co) teljes koncentrációjához viszonyított aránya. Az X-tengelya DNS kezdeti koncentrációjának termékének log-skálája (mól/liter)szorozva a reakció időtartamával (másodpercben)., Ennek az értéknek a jelölése kiságy, amelyet “kiságy” értéknek neveznek. Maga a görbe “kiságy” görbének nevezik. Mint látható, a görbe meglehetősen sima, amelyazt jelzi, hogy az újraindulás lassul, de fokozatosan egy idő alatt. Az egyik hasznos érték a Cot½, Akot érték, ahol a DNS fele újra be van állítva.
A DNS Denaturációs és Renaturációs kísérletekben részt vevő lépések
1. Nyírja a DNS-t körülbelül 400 bp méretre.
2. Denaturálja a DNS-t 100oC-ra melegítve.
3. Lassan hűtsük le, és különböző időközönként vegyünk mintát.
4., Határozza meg a % egyszálú DNS-t minden egyes időpontban.
egy adott fajra vonatkozó “kiságy” görbe alakja két tényező függvénye:
- a genom mérete vagy összetettsége; és
- az ismétlődő DNS mennyisége a genomban
Ha három faj, például a bakteriophagelambda,az E. coli és az élesztő genomjának “Cot½” görbéit ábrázoljuk, látni fogjuk, hogy azonos alakúak, de az élesztő Cot½-ja lesz a legnagyobb, az E. coli nextand lambda legkisebb., Fizikailag minél nagyobb a genomméret, annál hosszabb ideig tartmindegyik szekvenciának meg kell felelnie a kiegészítő szekvenciájának a megoldásban. Ez azért van, mert két egymást kiegészítő szekvenciának kell találkozniaminden más, mielőtt párosodnának. Minél összetettebb a genom, azaz minél több egyedi szekvencia áll rendelkezésre, annál hosszabb ideig tart, amíg bármelyik két egymást kiegészítő szekvencia találkozik egymással és párokkal. A hasonló oldatkoncentrációk miatt ez egy összetettebb fajt fog igénybe venni, amely már régóta eléri a Cot½-ot .,
ismétlődő DNS-szekvenciák, a faj genomjában többször megtalálható DNS-szekvenciák megkülönböztető hatással vannak a” Cot ” görbékre.Ha egy adott sorrendben képviselteti magát kétszer a genom lesz twocomplementary sorozatok párosítani, s mint ilyen lesz Gyerekágy érték felét mint nagy, mint egy sorozat képviseli, csak egyszer a genomban.
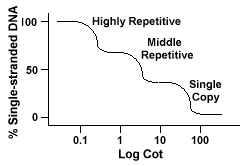
Az eukarióta genomoknak valójában számos olyan szekvenciájuk van, amelyeket az ismétlés különböző szintjein képviselnek., Egyetlen másolási szekvenciákat találnakvagy néhányszor a genomban. A functionalgeneket kódoló szekvenciák közül sok ebbe az osztályba tartozik. A középső ismétlődő DNS 10-1000-szer fordul elő a genomban. Ilyen például az rRNA és a trnagének, valamint a növényekben, például a kukoricában tárolt fehérjék. A középső dnakan 100-300 bp-ről 5000 bp-re változik, és a genomban szétszóródhat.A leggyakoribb szekvenciák találhatók a rendkívül ismétlődő DNAclass., Ezek a szekvenciák 100 000-1 millió alkalommal találhatók a genombanés mérete néhány-több száz bázis hosszú lehet. Ezek a kromoszóma olyan régióiban találhatók meg, mint a heterokromatin, a centromerek és a telomerek, és általában tandem ismétlésként vannak elrendezve. Thefollowing egy példa a tandemly ismételt szekvencia:
ATTATA ATTATA ATTATA / / ATTATA
Genomes, amelyek tartalmazzák ezeket a különböző osztályok szekvenciák reanneal egy differentmannner, mint genomes csak egy példányban szekvenciák., Ahelyett, hogy egy sima “kiságy” görbe lenne, három különálló görbe látható, amelyek mindegyike különböző ismétlési osztályt képvisel. A reanneal első szekvenciái a highlyrepetitive szekvenciák, mivel ezek sok példánya létezik a genomban, és mivel alacsony szekvencia-komplexitásuk van. A reanneális gén második része a középső repetitív DNS, a végső rész pedig az egypéldányos DNS., Az alábbi ábra egy “tipikus”eukarióta genom
a következő táblázat a kiválasztott fajok szekvenciaeloszlását mutatja.
| faj | szekvencia Eloszlás |
|---|---|
| baktériumok | 99.,27% – a Középső Ismétlődő 8% Erősen Ismétlődő |
| Kukorica | 30%, Egyetlen Példány 40% – a, Közel Ismétlődő 20% Erősen Ismétlődő |
| Búza | 10% – át Egyetlen Példány 83% – a Középső Ismétlődő 4% Erősen Ismétlődő |
| Osztályába | 55% – át Egyetlen Példány 27% – a Középső Ismétlődő 10% Erősen Ismétlődő |
Sorrend Interspersion
annak ellenére, hogy a genom szekvenciát jelent, a magasabb élőlények tartalmaz egyetlen példányt, middlerepetitive, többszörösen ismétlődő DNS szekvenciák, ezek a sorozatok nem arrangedsimilarly minden faj., A kiemelkedő elrendezést rövid időszaknak nevezikérdeklődés. Ezt az elrendezést ismételt szekvenciák jellemzik100-200 bp hosszúságú, 1000-2000bp hosszúságú egypéldányos szekvenciák között. Ez az elrendezés megtalálható állatokban, gombákban és növényekben.
a második típusú elrendezés hosszú távú interspersion. Ezt 5000 bp szakaszú ismétlődő szekvenciák jellemzik, amelyek 35 000 bp egypéldányos DNS-t kereszteznek. A Drosophila egy példaegy olyan fajból, amelynek ez a ritka szekvencia elrendezése van., Mindkét esetben ottismételt szekvenciák általában a középső ismétlődő osztályból származnak. Megbeszéltük, hol találhatók erősen ismétlődő szekvenciák.
eukarióta kromoszóma kariotípus
míg a baktériumok csak egyetlen kromoszómával rendelkeznek, az eukarióta Fajok legalább egy pár kromoszómával rendelkeznek. A legtöbbnek több párja van. Egy másik fontos szempont, hogy az eukarióta kromoszómákat csak a sejtosztódás során észlelik, nem pedig a sejtciklus minden szakaszában. Ezek a leginkábbkondenzált formában vannak a metafázis során,amikor a testvérkromatidokat csatolják.,Ez az elsődleges szakasz, amikor citogenetikai elemzést végeznek.
minden fajt kariotípus jellemez. A kariotípus ada kromoszómák számának leírása a normál diploid sejtben, valamintmint méreteloszlásuk. Például az emberi kromoszómának 23 párja van a kromoszómában, 22 szomatikus pár és egy pár nemi kromoszóma. A genetikai kutatás egyik fontos szempontja a kariotípus változásainak korrelálása az egyén fenotípusában bekövetkező változásokkal.
a genetika egyik fontos szempontja a kariotípus változásainak korrelálása a fenotípus változásaival., Például az emberek, akiknek extra 21-es kromoszómájuk Vanle Down-szindróma. Betoldások, törlések és a kromoszóma számának változásaa képzett citogenetikus detektálhatja, de ezeket a specifikus fenotípusokkal való korreláció nehéz.
a kariotípus kialakulásakor az első diszkrimináló paraméter a kromoszómák mérete és száma. Bár ez hasznos, nem biztosítelegendő részlet a szerkezet és a funkció (fenotípus) közötti korreláció kialakulásának megkezdéséhez., A kromoszómák közötti további megkülönböztetéshezolyan festékkel kezelik, amely reprodukálható módon festi a DNS-t. A festés után a régiók egy része enyhén foltos, mások pedig nehézkesek. Mint fentebb leírtuk, az enyhén festett régiókat nevezikeuchromatin, a sötét foltos régiót pedig hívjákheterochromatin. A választott jelenlegi festék a Giemsa folt, a kapott mintát g-sávos mintának nevezik.,
C-érték paradoxon
amellett,hogy leírja a szervezet genomját a kromoszómák számával, azt a DNS mennyisége is leírja egy haploid sejtben. Ez általában a haploid sejtenkénti DNS-mennyiség (általában pikogramokként kifejezve)vagy a haploid sejtenkénti kilobázisok száma, amelyet C-értéknek neveznek. Az eukarióta szervezetek egyik közvetlen jellemzője egy speciális anomáliát mutat, amelyet a molekuláris kutatás korai szakaszában fedeztek fel., Annak ellenére, hogy eukarióta szervezetekúgy tűnik, hogy 2-10-szer annyi gén van, mint a prokarióták, sok nagyságrenddel több DNS van a sejtben. Ezenkívül a DNS genomeisonkénti mennyisége nem korrelált egy faj feltételezett evolúciós összetettségével.Ezt a C érték paradoxonnak nevezik: a szervezet haploidcellájában lévő DNS mennyisége nem kapcsolódik evolúciós összetettségéhez. (Egy másik fontos szempont, amelyet szem előtt kell tartani, hogy nincs összefüggés a kromoszómák száma és az anorganizmus feltételezett evolúciós összetettsége között.,)
C Values of Organisms Used in Genetic Studies
| Species | Kilobases/haploid genome |
|---|---|
| E. coli |
4.5 x 103 |
| Human |
3.0 x 106 |
| Drosophila |
1.7 x 105 |
| Maize |
2.0 x 106 |
| Aribidopsis |
7.,0 x 104 |
A C értékek tartományának drámai példája látható a növény királyságbanahol az Arabidopsis az alsó végét és a liliomot (1,0 x 10^8 kb/haploidgenome) a komplexitás magas végét jelenti. Súly szempontjából ez 0,07 picogramsper haploid Arabidopsis genom és 100 pikogram / haploid lilygenome.
Genom-az egyetlen szülőtől örökölt kromoszómák teljes halmaza;az egyén teljes DNS-összetevője; a meghatározás gyakran kizárjaszervezetek