eukaryotisk kromosomstruktur
längden på DNA i kärnan är mycket större än storleken på den delningi vilken den finns. För att passa in i detta fack måste DNA varakondenserad på något sätt. Graden till vilken DNA kondenseras uttryckssom dess förpackningsförhållande.
packningsförhållande-längden av DNA dividerat med den längd i vilkenDet är förpackat
till exempel innehåller den kortaste humana kromosomen 4,6 x 107 bp DNA(ca 10 gånger genomets storlek av E. coli)., Detta motsvarar 14 000 µm utökat DNA. I sitt mest kondenserade tillstånd under mitos, denkromosom är ca 2 µm lång. Detta ger ett förpackningsförhållande på 7000 (14 000/2).
för att uppnå det totala förpackningsförhållandet förpackas DNA inte direkt i kromatin. I stället innehåller den flera hierarkier av organisation.Den första nivån av packning uppnås genom lindning av DNA runt ett proteincore för att producera en” pärlliknande ” struktur som kallas en nukleosom. Dettager ett förpackningsförhållande på ca 6., Denna struktur är invariant i både eukromatin och heterokromatin av alla kromosomer. Den andra nivån av packningär coiling av pärlor i en spiralstruktur som kallas 30 nm fibernsom finns i både interfas kromatin och mitotiska kromosomer. Dennastruktur ökar förpackningsförhållandet till ca 40. Den slutliga förpackningen uppstårnär fibern är organiserad i loopar, byggnadsställningar och domäner som ger ett slutligt förhållande på cirka 1000 i interfas kromosomer och cirka 10 000 inmitotiska kromosomer.,
eukaryota kromosomer består av ett DNA-proteinkomplex som är organiserat på ett kompakt sätt som gör det möjligt att lagra den stora mängden DNA i cellens kärna. Subenheten beteckning av kromosomen är kromatin.Den grundläggande enheten av kromatin är nukleosomen.,romatin – enheten för analys av kromosom; kromatin reflectsthe allmänna strukturen av kromosomen, men är inte unikt för någon particularchromosome
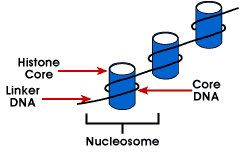
Nucleosome – enklaste förpackning struktur av DNA som finns inall eukaryota kromosomer; DNA är virad runt en octamer av små basicproteins som kallas histoner; 146 bp är lindade runt kärnan och remainingbases länk till nästa nucleosome; denna struktur medför negativa supercoiling
nucleosome består av cirka 200 bp lindade runt en histon octamerthat innehåller två kopior av histonproteiner H2A, H2B, H3 och H4., Dessa är kända som kärnhistoner. Histoner är grundläggande proteiner som har en affinitetför DNA och är de mest rikliga proteinerna i samband med DNA. Aminoacidsekvensen för dessa fyra histoner bevaras vilket tyder på en liknande funktion för alla.
längden på DNA som är associerad med nukleosomenheten varierar mellanspecies. Men oavsett storlek är två DNA-komponenter inblandade. CoreDNA är DNA som faktiskt är associerad med histone octamer.Detta värde är invariant och är 146 baspar., Kärnan DNA bildar två loopsaround octamer, och detta tillåter två regioner som är 80 bp isär tobe förs i närheten. Således kan två sekvenser som är långt ifrån varandrainteragera med samma reglerande protein för att kontrollera genuttryck. TheDNA som är mellan varje histon octamer kallas länkaren DNAand kan variera i längd från 8 till 114 baspar. Denna variation är speciellspecifik, men variation i linker-DNA-längd har också associerats med organismens utvecklingsstadium eller specifika regioner i genomet.,
nästa organisationsnivå för kromatin är 30 nm-fibern. Dettavisas vara en solenoidstruktur med cirka 6 nukleosomer per tur. Dettager ett förpackningsförhållande på 40, vilket innebär att varje 1 µm längs axisinnehåller 40 µm DNA. Stabiliteten hos denna struktur krävernärvaro av den sista medlemmen av histongenfamiljen, Histon H1., Eftersomupplevelser som avlägsnar H1 från kromatin upprätthåller nukleosomen, men inte den 30 nm-strukturen, drogs slutsatsen att H1 är viktigt för stabiliseringen av 30 nm-strukturen.
den slutliga förpackningsnivån kännetecknas av 700 nm-strukturen seenin metafaskromosomen. Den kondenserade delen av kromatin har acharacteristic byggnadsställningar struktur som kan detekteras i metafasekromosomer. Detta verkar vara resultatet av omfattande looping av dnain kromosomen.
de sista definitionerna som måste presenteras är euchromatin ochheterochromatin., När kromosomer färgas med färgämnen verkar de haväxla lätt och mörkt färgade områden. De lättfärgade regionerna är euchromatin och innehåller enkopiering, genetiskt aktivt DNA. De mörkfärgade regionerna är heterokromatin och innehåller repetitiva sekvenser som är genetiskt inaktiva.
centromerer och telomerer
centromerer och telomerer är två väsentliga egenskaper hos alla eukaryota kromosomer. Var och en ger en unik funktion som är absolut nödvändig för kromosomens stabilitet., Centromeres krävs för segregering av centromeren under meios och mitos, och teleomerer ger terminal stabilitet mot kromosomen och säkerställa dess överlevnad.
centromerer är de kondenserade regionerna inom kromosomen som är ansvariga för den exakta segregeringen av de replikerade kromosomerna under mitos och meios. När kromosomer färgas visar de vanligtvis en mörkfärgad region som är centromeren. Under mitos måste centromeren som delas av systerkromatiderna dela upp så att kromatiderna kan migrera tillmotsatta poler i cellen., Å andra sidan, under den första meiotiska divisionencentromeren av systerkromatider måste förbli intakta, medan de under meiosisII måste fungera som de gör under mitos. Därför är centromeren enviktig komponent i kromosomstruktur och segregering.
inom centromereregionen har de flesta arter flera platser där spindelfibrer fäster, och dessa platser består av DNA såväl som protein. Den faktiska platsen där fästet inträffar kallas kinetochore och består av både DNA och protein. DNA-sekvensen inom dessa regioner kallascen-DNA., Eftersom CEN-DNA kan flyttas från en kromosom till en annan och fortfarande ger kromosomen förmågan att segregera, får dessa sekvenser inte ge någon annan funktion.
vanligtvis CEN DNA är ca 120 baspar lång och består av flera underdomäner, CDE-i, CDE-II och CDE-III. mutationer i de två första underdomänerna har ingen effekt på segregation, men en punktmutation i CDE-III underdomänen eliminerar helt centromerens förmåga att fungera under kromosomsegregering., Därför måste CDE-III vara aktivt involverad i bindningen av spindelfibrerna till centromeren.
proteinkomponenten i kinetochore karakteriseras först nu. Ett komplex av tre proteiner som kallas Cbf-III binder till normala CDE-III regioner men kan inte binda till en CDE-III-region med en punktmutation som förhindrar mitotisk segregering. Vidare eliminerar mutanter av generna som kodar för Cbf-III-proteinerna också förmågan för kromosomer att segregera under mitos., Ytterligare analyser av DNA-och proteinkomponenterna i centromeren är nödvändiga för att fullt ut förstå mekaniken för kromosomsegregering.
telomerer är regionen av DNA i slutet av den linjära eukaryotikkromosom som krävs för replikering och stabilitet av kromosomen.McClintock kände igen sina speciella egenskaper när hon märkte, att om tvåkromosomer bröts i en cell, kunde slutet av en fästa vid den andraoch vice versa. Vad hon aldrig observerade var brokenendens fastsättning till slutet av en obruten kromosom., Således ändarna av brutna kromosomerär klibbiga, medan den normala änden inte är klibbig, vilket tyder på ändarna avkromosomer har unika egenskaper. Vanligtvis, men inte alltid, är telomericDNA heterokromatisk och innehåller direkta tandemly upprepade sekvenser. Följande tabell visar upprepningssekvenserna för flera arter. Dessa är ofta av formen (T/A)xGy där x är mellan 1 och 4 och y är större än 1.,
Telomere Repeat Sequences
| Species | Repeat Sequence |
|---|---|
| Arabidopsis | TTTAGGG |
| Human | TTAGGG |
| Oxytricha | TTTTGGGG |
| Slime Mold | TAGGG |
| Tetrahymena | TTGGGG |
| Trypanosome | TAGGG |
| Yeast | (TG)1-3TG2-3 |
Notice that the number of TG sequences and the number of cytosines in the yeast sequence varies., Åtminstone för jäst har det visat sig att olika stammar innehåller olika längder av teleomerer och att längden är under genetisk kontroll.
den primära svårigheten med telomerer är replikeringen av den släpande strängen.Eftersom DNA-syntes kräver en RNA-mall (som ger den fria 3 ’ – OHgroup) för att Primera DNA-replikering, och denna mall försämras så småningom, skulle en kort enkelsträngad region lämnas i slutet av kromosomen. Denna region skulle vara mottaglig för enzymer som försämrar enkelsträngat DNA., Resultatet skulle vara att kromosomens längd skulle förkortas efter varje uppdelning. Men detta ses inte.
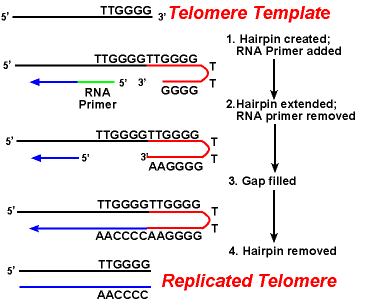
telomeras-enzymernas verkan säkerställer att ändarna av de släpande strängarna replikeras korrekt. Ett väl studerat system involverar Tetrahymena protozoa organismen. Telomererna i denna organism slutar i sekvensen 5′-TTGGGG-3′. Telomeras lägger till en serie av 5′-TTGGGG-3 ’ upprepningar till ändarna av den släpande strängen. En hårnål uppstår när ovanliga baspar mellan guaninrester i upprepningsformen., Därefter avlägsnas rna-primern, och 5-änden av den fördröjda strängen kan användas för DNA-syntes. Ligering sker mellan den färdiga släpande strängen och hårnålen. Slutligen avlägsnas hårnålen vid 5’ – ttgggg-3 ’ – upprepningen. Således replikeras änden av kromosomen troget. Följande figur visar dessa steg.,
replikeringen av telomerer
analys av DNA-sekvenser i eukaryota genomer
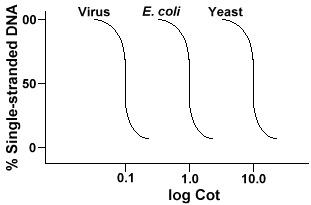
den teknik som används för att bestämma sekvensens komplexitet innebär denaturering och omräkning av DNA. DNA denaturedby uppvärmning som smälter H-bindningar och gör DNA enkelsträngade. Om DNA kyls snabbt, förblir DNA enkelsträngad. Men om DNAis får svalna långsamt, kommer sekvenser som kompletterar varandra att hitta varandra och så småningom basparet igen., Den kurs till vilken DNA reanneals(en annan term för renature) är en funktion av den art från vilken DNAwas isolerade. Nedan är en kurva som erhålls från ett enkelt genom.
y-axeln är procentandelen av DNA som förblir enkelsträngad. Detta uttryckas som ett förhållande mellan koncentrationen av enkelsträngat DNA (C)och den totala koncentrationen av Start-DNA (Co). X-axeln ären log-skala av produkten av den ursprungliga koncentrationen av DNA (i mol/liter)multiplicerad med tiden reaktionen fortsatte (i sekunder)., Designationenför detta värde är barnsäng och kallas ”barnsäng” – värdet. Kurvan självkallas en” Cot ” – kurva. Som kan ses är kurvan ganska smidig somindikerar att reannealing sker sakta men gradvis under en period avtid. Ett särskilt värde som är användbar är Cot½ , theCot värde där hälften av DNA har reannealed.
steg involverade i DNA-denaturering och Renatureringsexperiment
1. Klippa DNA till en storlek av ca 400 bp.
2. Denature DNA genom uppvärmning till 100oC.
3. Kyl långsamt och ta prover med olika tidsintervall.
4., Bestäm % enkelsträngat DNA vid varje tidpunkt.
formen på en ”Cot” – kurva för en viss art är en funktion av två faktorer:
- genomets storlek eller komplexitet; och
- mängden repetitivt DNA inom genomet
om vi plottar ”Cot” – kurvorna för genomet av tre arter som bakteriofagelambda, E. coli och jäst kommer vi att se att de har samma form,men Cot½ av jästen kommer att vara störst, och att E. coli nextand Lambda minsta., Fysiskt, ju större genomstorleken desto längre är detkommer att ta för någon sekvens att stöta på dess komplementära sekvens ilösningen. Detta beror på att två kompletterande sekvenser måste stöta påvarje annan innan de kan para sig. Ju mer komplexa genomet, det vill sägamer unika sekvenser som är tillgängliga, desto längre kommer det att ta för någotvå kompletterande sekvenser att stöta på varandra och par. Med tanke på liknandekoncentrationer i lösningen kommer det att ta en mer komplex art lång tid att nå Cot½ .,
upprepade DNA-sekvenser, DNA-sekvenser som finns mer än en gång i artens Genom har särskiljande effekter på ”Cot” – kurvorna.Om en specifik sekvens representeras två gånger i genomet kommer det att ha tvåkomplementära sekvenser att para med och som sådan kommer att ha ETT barnsäng värde halvas stor som en sekvens som representeras endast en gång i genomet.
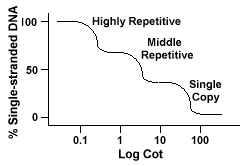
eukaryota genom har faktiskt ett brett spektrum av sekvenser som representeras på olika nivåer av upprepning., Enstaka kopieringssekvenser finns en gångeller några gånger i genomet. Många av de sekvenser som kodar functionalgenes faller i denna klass. Mellan repetitivt DNA finns från 10s -1000 gånger i genomet. Exempel på dessa skulle vara rRNA och trnagener och lagringsproteiner i växter som majs. Mellersta repetitiva dnacan varierar från 100-300 bp till 5000 bp och kan spridas genom hela genomet.De mest rikliga sekvenserna finns i den mycket repetitiva DNAclass., Dessa sekvenser finns från 100 000 till 1 miljon gånger i genometoch kan variera i storlek från några till flera hundra baser i längd. Dessa skillnader finns i kromosomregioner som heterochromatin,centromeres och telomerer och tenderar att ordnas som en tandemrepetitioner. Thefollowing är ett exempel på en tandemly upprepad sekvens:
ATTATA ATTATA ATTATA // ATTATA
Genom att innehålla dessa olika klasser av sekvenser reanneal i en differentmanner än arvsmassa med endast en enda kopiera sekvenser., I stället för att ha asingle slät” Cot ” kurva, kan tre distinkta kurvor ses, var och en representeraren annan repetitionsklass. De första sekvenserna att reanneal är highlyrepetitive sekvenser eftersom så många kopior av dem finns i genomet, och eftersom de har en låg sekvens komplexitet. Den andra delen av thegenome att reanneal är mitt repetitivt DNA, och den sista delen toreanneal är den enda kopiera DNA., Följande diagram visar kurvan”spjälsäng ”för ett” typiskt”eukaryotiskt genom
Följande tabell ger sekvensfördelningen för utvalda arter.
| arter | Sekvensfördelning |
|---|---|
| bakterier | 99.,27% Middle Repetitive 8% Highly Repetitive |
| majs | 30% Single Copy 40% Middle Repetitive 20% Highly Repetitive |
| vete | 10% Single Copy 83% Middle Repetitive 4% Highly Repetitive |
| Arabidopsis | 55% Single Copy 27% Middle repetitive 10% highly repetitive |
sequence Interspersion
Även om genomen av högre organismer innehåller enstaka kopior, mittenrepetitiva och mycket repetitiva DNA-sekvenser, är dessa sekvenser inte ordnade på samma sätt i alla arter., Det framträdande arrangemanget kallas kort periodinterspersion. Detta arrangemang kännetecknas av upprepade sekvenser100-200 bp i längd interspersed bland enstaka kopieringssekvenser som är 1000-2000bp i längd. Detta arrangemang finns i djur, svampar och växter.
den andra typen av arrangemang är lång period interspersion. Detta kännetecknas av 5000 bp-sträckor av upprepade sekvenser som varvasinom regioner med 35 000 bp av ENKELKOPIERAD DNA. Drosophila är ett exempelav en art med detta ovanliga sekvensarrangemang., I båda fallen är dessa sekvenser vanligtvis från den mellersta repetitiva klassen. Vi diskuteradeovan där mycket repetitiva sekvenser finns.
eukaryotisk kromosom karyotyp
bakterier har endast en enda kromosom, eukaryota arter har minst ett par kromosomer. De flesta har mer än ett par. En annan relevant punkt är att eukaryotiska kromosomer detekteras endast under celldivision och inte under alla stadier av cellcykeln. De är i sin mestkondenserad form under metafas när systerkromatiderna är fästa.,Detta är det primära steget när cytogenetisk analys utförs.
varje art kännetecknas av en karyotyp. Karyotypen är enbeskrivning av antalet kromosomer i den normala diploida cellen, liksom deras storleksfördelning. Till exempel har den mänskliga kromosomen 23 parav kromosom, 22 somatiska Par och ett par sexkromosomer. En viktigaspekt av genetisk forskning korrelerar förändringar i karyotypen med förändringari individens fenotyp.
en viktig aspekt av genetiken är att korrelera förändringar i karyotyp medförändringar i fenotyp., Till exempel människor som har en extra kromosom 21har Downs syndrom. Infogningar, deletioner och förändringar i kromosomnummerkan detekteras av den skickliga cytogeneticisten, men det är svårt att korrelera dessa medspecifika fenotyper.
den första diskriminerande parametern när man utvecklar en karyotyp är storlekenoch antalet kromosomer. Även om detta är användbart, ger det intetillräcklig detalj för att börja utvecklingen av en korrelation mellan struktureroch funktion (fenotyp)., För att ytterligare skilja mellan kromosomer, debehandlas med ett färgämne som fläckar DNA på ett reproducerbart sätt. Efter det att vissa av regionerna är lätt färgade och andra är tungahållna. Som beskrivits ovan kallas de lättfärgade regionernaukromatin, och den mörka färgade regionen kallasheterochromatin. Det nuvarande färgämnet av chose är Giemsa-fläcken, och det resulterande mönstret kallas G-bandingmönstret.,
c-värde Paradox
förutom att beskriva genomet hos en organism med dess antal kromosomer beskrivs det också av mängden DNA i en haploidcell. Detta är vanligtvisuttryckt som mängden DNA per haploidcell (vanligtvis uttryckt som picogram)eller antalet kilobaser per haploidcell och kallas C-värdet. Ettomedelbart inslag i eukaryota organismer belyser en specifik anomali som upptäcktes tidigt i molekylär forskning., Även om eukaryotiska organismerförekommer att ha 2-10 gånger så många gener som prokaryoter, har de många ordrar av storleksordning mer DNA i cellen. Vidare korreleras mängden DNA per Genom inte med den förmodade evolutionära komplexiteten hos en art.Detta anges som C-värdeparadoxen: mängden DNA i en organisms haploidcell är inte relaterad till dess evolutionära komplexitet. (En annan viktig punkt att komma ihåg är att det inte finns något samband mellanantalet kromosomer och den förmodade evolutionära komplexiteten hos en organism.,)
C Values of Organisms Used in Genetic Studies
| Species | Kilobases/haploid genome |
|---|---|
| E. coli |
4.5 x 103 |
| Human |
3.0 x 106 |
| Drosophila |
1.7 x 105 |
| Maize |
2.0 x 106 |
| Aribidopsis |
7.,0 x 104 |
Ett dramatiskt exempel på utbudet av C-värden kan ses i anläggningen kingdomwhere Arabidopsis representerar den lägre änden och lily (1,0 x 10^8 kb/haploidgenome) den övre delen av komplexitet. I vikt termer är detta 0.07 picogramsper haploida Arabidopsis genomet och 100 picogram per haploida lilygenome.
genom – den kompletta uppsättningen kromosomer som ärvs från en enda förälder; den fullständiga DNA-komponenten hos en individ; definitionen utesluter oftaorganeller