eukaryotische Chromosoomstructuur
De lengte van DNA in de kern is veel groter dan de grootte van het compartiment waarin het zit. Om in dit compartiment te passen moet het DNA op een of andere manier gecondenseerd worden. De mate waarin DNA wordt gecondenseerd wordt uitgedrukt als zijn verpakkingsverhouding.
Verpakkingsverhouding-de lengte van DNA gedeeld door de lengte waarin het verpakt is
het kortste menselijke chromosoom bevat bijvoorbeeld 4,6 x 107 bp DNA(ongeveer 10 keer de genoomgrootte van E. coli)., Dit komt overeen met 14.000 µm van uitgebreid DNA. In zijn meest gecondenseerde staat tijdens mitose, is hetchromosoom ongeveer 2 µm lang. Dit geeft een verpakkingsverhouding van 7000 (14.000 / 2).
om de totale verpakkingsverhouding te bereiken, wordt DNA niet rechtstreeks verpakt in de uiteindelijke structuur van chromatine. In plaats daarvan bevat het verschillende hiërarchieën van de organisatie.Het eerste niveau van verpakking wordt bereikt door het wikkelen van DNA rond proteincore om een “parel-als” structuur genoemd nucleosome te produceren. Dit geeft een verpakkingsverhouding van ongeveer 6., Deze structuur is invariant in zowel theeuchromatin als heterochromatin van alle chromosomen. Het tweede niveau van het verpakken is het rollen van parels in een spiraalvormige structuur genoemd 30 nm fiberdie in zowel interphase chromatin als mitotic chromosomen wordt gevonden. Deze structuur verhoogt de verpakkingsverhouding tot ongeveer 40. De definitieve verpakking komt voor wanneer de vezel in lussen, steigers en domeinen wordt georganiseerd die een definitieve verpakkingsverhouding van ongeveer 1000 in interfasechromosomen en ongeveer 10.000 inmitotic chromosomen geven.,
eukaryotische chromosomen bestaan uit een DNA-eiwitcomplex dat op een compacte manier is georganiseerd waardoor de grote hoeveelheid DNA in de celkern kan worden opgeslagen. De subeenheid aanduiding van het chromosoom is chromatine.De fundamentele eenheid van chromatine is het nucleosoom.,romatin – de eenheid van analyse van het chromosoom; chromatine reflectsthe algemene structuur van het chromosoom, maar is niet uniek voor elke particularchromosome
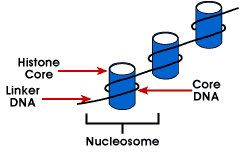
Nucleosome – eenvoudigste verpakking structuur van het DNA dat is gevonden inall eukaryotische chromosomen; DNA is verpakt rond een octamer van kleine basicproteins genoemd histonen; 146 bp is gewikkeld rond de kern en de remainingbases link naar de volgende nucleosome; deze structuur veroorzaakt een negatieve supercoiling
De nucleosome bestaat uit ongeveer 200 bp gewikkeld rond een histon octamerthat bevat twee exemplaren van de histon-eiwitten H2A, H2B, H3 en H4., Deze staan bekend als de core histones. Histonen zijn basisproteã nen die een affiniteit voor DNA hebben en zijn de overvloedigste proteã nen verbonden aan DNA. De aminozuurvolgorde van deze vier histonen wordt behouden die een gelijkaardige functie voor allen suggereren.
De lengte van DNA die geassocieerd is met de nucleosoom-eenheid varieert tussen soorten. Maar ongeacht de grootte zijn er twee DNA-componenten bij betrokken. CoreDNA is het DNA dat eigenlijk wordt geassocieerd met de Histon octamer.Deze waarde is invariant en is 146 basisparen., De kern DNA vormt twee lijnen rond de octamer, en dit laat twee gebieden toe die 80 BP apart zijn om in dichte nabijheid te worden gebracht. Aldus, kunnen twee opeenvolgingen die ver uit elkaar zijn met dezelfde regelgevende proteã ne interacteren om genuitdrukking te controleren. De DNA die tussen elke histone octamer is wordt genoemd linker DNA en kan in lengte van 8 tot 114 basisparen variëren. Deze variatie is soortspecifiek, maar variatie in de lengte van linkerdna is ook geassocieerd met het ontwikkelingsstadium van het organisme of specifieke gebieden van het genoom.,
het volgende niveau van organisatie van het chromatine is de 30 nm vezel. Dit lijkt een solenoïde structuur met ongeveer 6 nucleosomen per beurt. Dit geeft een verpakkingsverhouding van 40, wat betekent dat elke 1 µm langs de as 40 µm van DNA bevat. De stabiliteit van deze structuur vereist de aanwezigheid van het laatste lid van de histongenfamilie, Histon H1., Omdat de stoffen die H1 uit chromatine verwijderen het nucleosoom behouden, maar niet de 30 nm-structuur, werd geconcludeerd dat H1 belangrijk is voor de stabilisering van de 30 nm-structuur.
het uiteindelijke verpakkingsniveau wordt gekenmerkt door de 700 nm-structuur in het metafasechromosoom. Het gecondenseerde stuk chromatin heeft een kenmerkende steigerstructuur die in metafasechromosomes kan worden ontdekt. Dit schijnt om het resultaat van uitgebreide lus van DNAin het chromosoom te zijn.
de laatste definities die moeten worden gepresenteerd zijn euchromatine enheterochromatine., Wanneer chromosomen met kleurstoffen worden bevlekt, lijken zij licht en donker gekleurd gebieden te hebben. De licht bevlekte gebieden zijn euchromatin en bevatten enig-exemplaar, genetisch actief DNA. De donker gekleurde gebieden zijn heterochromatine en bevatten repetitieve opeenvolgingen die genetisch inactief zijn.
centromeren en telomeren
centromeren en telomeren zijn twee essentiële kenmerken van alle eukaryotische chromosomen. Elk biedt een unieke functie die absoluut noodzakelijk is voor de stabiliteit van het chromosoom., Centromeren worden vereist voor de scheiding van centromeer tijdens meiosis en mitose, en teleomeren verstrekken eindstabiliteit aan het chromosoom en verzekeren zijn overleving.
centromeren zijn de gecondenseerde gebieden binnen het chromosoom die verantwoordelijk zijn voor de nauwkeurige segregatie van de gerepliceerde chromosomen tijdens mitose en meiose. Wanneer chromosomen worden bevlekt tonen zij typisch een donker-bevlekt gebied dat centromeer is. Tijdens mitose, moet centromeer dat door zusterchromatiden wordt gedeeld zo verdelen dat chromatiden ook kunnen migrerenposite polen van de cel., Aan de andere kant, tijdens de eerste meiotische verdeling moet het centromeer van zusterchromatiden intact blijven, terwijl ze tijdens meiosisII moeten handelen zoals ze doen tijdens mitose. Daarom is centromeer een belangrijke component van chromosoomstructuur en segregatie.
binnen het centromeergebied hebben de meeste soorten verschillende plaatsen waar spindelvezels zich hechten, en deze plaatsen bestaan uit zowel DNA als eiwit. De daadwerkelijke plaats waar de gehechtheid voorkomt wordt genoemd kinetochore en uit zowel DNA als proteã ne samengesteld. De opeenvolging van DNA binnen deze gebieden wordt genoemd cen DNA., Omdat CEN-DNA van het ene chromosoom naar het andere kan worden verplaatst en het chromosoom nog steeds van de capaciteit kan voorzien om te scheiden, moeten deze opeenvolgingen geen andere functie verstrekken.
meestal is CEN-DNA ongeveer 120 basenparen lang en bestaat uit verschillende subdomeinen, CDE-I, CDE-II en CDE-III. mutaties in de eerste twee subdomeinen hebben geen effect op segregatie, maar een puntmutatie in het CDE-III subdomein elimineert volledig het vermogen van de centromeer om te functioneren tijdens chromosoomsegregatie., Daarom moet CDE-III actief betrokken zijn bij de binding van de spindelvezels aan de centromeer.
de eiwitcomponent van de kinetochoor wordt nu pas gekarakteriseerd. Een complex van drie proteã nen genoemd Cbf-III bindt aan normale CDE-III gebieden maar kan niet aan een CDE-III gebied met een puntmutatie binden die mitotic segregatie verhindert. Bovendien elimineren mutanten van de genen die de Cbf-III-eiwitten coderen ook het vermogen voor chromosomen om te scheiden tijdens mitose., Aanvullende analyses van de DNA-en eiwitcomponenten van centromeer zijn noodzakelijk om de mechanica van chromosoomscheiding volledig te begrijpen.
telomeren zijn het gebied van DNA aan het einde van het lineaire eukaryoticchromosoom dat nodig is voor de replicatie en stabiliteit van het chromosoom.McClintock herkende hun speciale kenmerken toen ze merkte, dat als tweechromosomen werden gebroken in een cel, het einde van de ene kon hechten aan de andere en vice versa. Wat ze nooit zag was de gehechtheid van de brokenend aan het einde van een ongebroken chromosoom., Aldus zijn de einden van gebroken chromosomen kleverig, terwijl het normale eind niet kleverig is, die de einden van chromosomes voorstellen unieke eigenschappen hebben. Gewoonlijk, maar niet altijd, is telomericDNA heterochromatisch en bevat directe tandemly herhaalde opeenvolgingen. De volgende tabel toont de herhaalsequenties van verschillende soorten. Deze zijn vaak van de vorm (T/A)xGy waarbij x tussen 1 en 4 ligt en y groter is dan 1.,
Telomere Repeat Sequences
| Species | Repeat Sequence |
|---|---|
| Arabidopsis | TTTAGGG |
| Human | TTAGGG |
| Oxytricha | TTTTGGGG |
| Slime Mold | TAGGG |
| Tetrahymena | TTGGGG |
| Trypanosome | TAGGG |
| Yeast | (TG)1-3TG2-3 |
Notice that the number of TG sequences and the number of cytosines in the yeast sequence varies., Ten minste voor gist is aangetoond dat verschillende stammen verschillende lengtes teleomeren bevatten en dat de lengte onder genetische controle is.
het primaire probleem met telomeren is de replicatie van het achterblijvende onderdeel.Omdat de synthese van DNA een malplaatje van RNA vereist (dat de vrije 3′-Ohgroep verstrekt) om de replicatie van DNA te prime, en dit malplaatje uiteindelijk wordt gedegradeerd, zou een kort single-stranded gebied aan het eind van het chromosoom worden verlaten. Dit gebied zou vatbaar zijn voor enzymen die single-stranded DNA degraderen., Het resultaat zou zijn dat de lengte van het chromosoom na elke verdeling zou worden verkort. Maar dit wordt niet gezien.
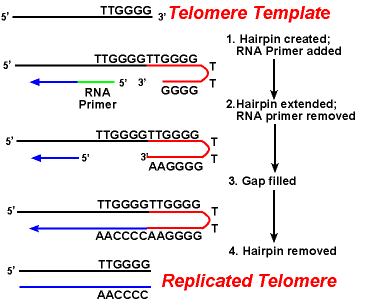
de werking van de telomerase-enzymen zorgt ervoor dat de uiteinden van de achterblijvende strengen correct worden gerepliceerd. Een goed bestudeerd systeem omvat het Tetrahymena protozoa organisme. De telomeren van dit organisme eindigen in de opeenvolging 5′-TTGGGG-3′. Telomerase voegt een reeks 5′ – ttgggg-3 ‘ herhaalt aan de einden van de achterblijvende bundel toe. Een haarspeld treedt op wanneer ongebruikelijke base paren tussen guanine residuen in de herhaalvorm., Vervolgens wordt de inleiding van RNA verwijderd, en het einde 5′ van de achterblijvende bundel kan voor de synthese van DNA worden gebruikt. Ligatie vindt plaats tussen de afgewerkte achterblijvende streng en de haarspeld. Tot slot wordt de haarspeld verwijderd bij de 5’-TTGGGG-3 ‘ herhaling. Aldus wordt het eind van het chromosoom getrouw herhaald. De volgende figuur toont deze stappen.,
de replicatie van telomeren
analyse van DNA-sequenties in eukaryotische genomen
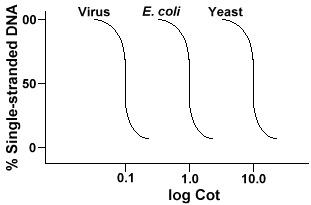
de techniek die wordt gebruikt om de sequentiecomplexiteit van een genomein te bepalen, omvat de denaturatie en renaturatie van DNA. DNA wordt gedenatureerd door het verwarmen dat de H-banden smelt en DNA single-stranded maakt. Als het DNA snel wordt gekoeld, blijft het DNA single-stranded. Maar als de DNAis toegestaan om langzaam af te koelen, sequenties die complementair zijn zullen elkaar vinden en uiteindelijk base paar weer., De snelheid waarmee de DNA reanneals (een andere term voor renature) is een functie van de species waarvan de Dnawais geà soleerd. Hieronder is een kromme die uit een eenvoudig genoom wordt verkregen.
De Y-as is het percentage van het DNA dat single stranded blijft. Dit wordt uitgedrukt als Verhouding van de concentratie van single-stranded DNA (C)tot de totale concentratie van het begin van DNA (Co). De x-as iseen log-schaal van het product van de initiële concentratie van DNA (in mol/liter)vermenigvuldigd met de duur van de reactie (in seconden)., De aanduiding voor deze waarde is Cot en wordt de “Cot” waarde genoemd. De curve zelf wordt een “Cot” curve genoemd. Zoals kan worden gezien is de curve vrij glad, wat aangeeft dat het opnieuw gloeien plaatsvindt vertragen maar geleidelijk over een periode van tijd. Één bepaalde waarde die nuttig is is Cot½, theCot-waarde waar de helft van DNA opnieuw is geopend.
stappen betrokken bij DNA-denaturatie-en Renaturatieexperimenten
1. Schuin het DNA tot een grootte van ongeveer 400 bp.
2. Denature het DNA door te verwarmen tot 100oC.
3. Koel langzaam af en neem monsters op verschillende tijdstippen.
4., Bepaal de % single-stranded DNA op elk tijdstip.
de vorm van een “Cot” – curve voor een bepaalde soort is een functie van twee factoren:
- De grootte of complexiteit van het genoom; en
- de hoeveelheid repetitief DNA in het genoom
als we de “Cot” – curves van het genoom van drie soorten zoals bacteriofagelambda, E. coli en gist plotten,zullen we zien dat ze dezelfde vorm hebben, maar de Cot½ van de gist zal het grootst zijn, E. coli Next en lambda kleinste., Fysiek, hoe groter de genoomgrootte hoe langer het zal duren voor om het even welke opeenvolging om zijn complementaire opeenvolging in de oplossing te ontmoeten. Dit is omdat twee complementaire opeenvolgingen elkaar moeten ontmoeten voordat zij kunnen paren. Hoe complexer het genoom, dat de meer unieke opeenvolgingen is die beschikbaar zijn, hoe langer het zal duren voor om het even welke twee complementaire opeenvolgingen elkaar en paar ontmoeten. Gegeven vergelijkbare concentraties in oplossing, zal een complexere soort langer nodig hebben om Cot½ te bereiken .,
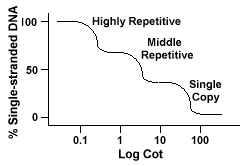
herhaalde DNA-sequenties, DNA-sequenties die meer dan eens in het genoom van de soort worden aangetroffen, hebben een onderscheidend effect op de krommen van” Cot”.Als een specifieke opeenvolging tweemaal in het genoom wordt vertegenwoordigd zal het twee complementaire opeenvolgingen hebben om met te paren en als zodanig zal een Cot-waarde halfas groot hebben aangezien een opeenvolging slechts eenmaal in het genoom wordt vertegenwoordigd.
eukaryotische genomen hebben eigenlijk een brede reeks sequenties die op verschillende niveaus van herhaling worden vertegenwoordigd., De enige opeenvolgingen van het exemplaar worden gevonden onceof een paar keer in het genoom. Veel van de opeenvolgingen die functionalgenes coderen vallen in deze klasse. Het middelste repetitieve DNA wordt gevonden van 10s -1000 keer in het genoom. Voorbeelden hiervan zouden rRNA en tRNAgenes en opslagproteã nen in installaties zoals maïs omvatten. Midden repetitieve DNAcan variëren van 100-300 BP tot 5000 bp en kan door het genoom worden verspreid.De meest voorkomende opeenvolgingen worden gevonden in de hoogst repetitieve DNAclass., Deze opeenvolgingen worden gevonden van 100.000 tot 1 miljoen keer in het genoom en kunnen in grootte van een paar aan verscheidene honderd basissen in lengte variëren. Deze gevolgen worden gevonden in gebieden van het chromosoom zoals heterochromatin,centromeres en telomeren en neigen om als tandem herhalingen worden geschikt. Het volgende is een voorbeeld van een tandemly herhaalde sequentie:
ATTATA ATTATA ATTATA // ATTATA
genomen die deze verschillende klassen van sequenties bevatten, worden heranneal in een andere manier dan genomen met slechts één kopie sequenties., In plaats van een enkele gladde “Cot” curve, drie verschillende curves kunnen worden gezien, elk vertegenwoordigen een verschillende herhaling klasse. De eerste opeenvolgingen om opnieuw te worden geannexeerd zijn de zeer repetitieve opeenvolgingen omdat er zoveel exemplaren van hen in het genoom bestaan,en omdat zij een lage opeenvolgingscomplexiteit hebben. Het tweede gedeelte van thegenome aan reanneal is het midden repetitieve DNA, en het definitieve gedeelte om te reanneal is het enige exemplaar DNA., Het volgende diagram toont de”Cot” – curve voor een” typisch”eukaryotisch genoom
de volgende tabel geeft de sequentieverdeling voor geselecteerde soorten weer.
| Species | Sequentiedistributie | bacteriën | 99.,27% Midden Repetitieve 8% Sterk Repetitieve |
|---|---|
| Maïs | 30% Één Kopie 40% Midden Repetitieve 20% Sterk Repetitieve |
| Tarwe | 10% Één Kopie 83% Midden Repetitieve 4% Uiterst Repetitieve |
| Arabidopsis | 55% Één Kopie 27% Midden Repetitieve 10% Sterk Repetitieve |
Volgorde Interspersion
hoewel het genoom van hogere organismen bevatten enkel exemplaar, middlerepetitive en uiterst repetitieve DNA sequenties, deze reeksen zijn niet arrangedsimilarly in alle soorten., De prominente arrangement heet korte periodeinterspersie. Deze regeling wordt gekenmerkt door herhaalde opeenvolgingen 100-200 bp in lengte die tussen enige exemplaaropeenvolgingen worden afgewisseld die 1000-2000bp in lengte zijn. Deze regeling is te vinden in dieren, schimmels en planten.
het tweede type arrangement is interpersieperiode voor lange perioden. Dit wordt gekenmerkt door 5000 BP strekken van herhaalde opeenvolgingen die binnen gebieden van 35.000 bp van enig exemplaardna worden afgewisseld. Drosophila is een voorbeeld van een soort met deze ongewone volgorde regeling., In beide gevallen, therepeated opeenvolgingen zijn gewoonlijk van de Midden repetitieve klasse. We discussieerden over waar zeer repetitieve sequenties worden gevonden.
eukaryotisch Chromosoomkaryotype
terwijl bacteriën slechts één chromosoom hebben, hebben eukaryotische soorten ten minste één paar chromosomen. De meeste hebben meer dan één paar. Een ander relevant punt is dat eukaryotic chromosomen worden ontdekt slechts voorkomen tijdens celldivision en niet tijdens alle stadia van de celcyclus. Ze zijn in hun meest gecondenseerde vorm tijdens metafase wanneer de zusterchromatiden zijn bevestigd.,Dit is het primaire stadium wanneer cytogenetische analyse wordt uitgevoerd.
elke soort wordt gekenmerkt door een karyotype. Het karyotype is een beschrijving van het aantal chromosomen in de normale diploïde cel, evenals hun grootteverdeling. Bijvoorbeeld, heeft het menselijke chromosoom 23 paar chromosoom, 22 somatische paren en één paar geslachtschromosomen. Een belangrijk aspect van genetisch onderzoek correleert veranderingen in het karyotype met veranderingen in het fenotype van het individu.
een belangrijk aspect van genetica is het correleren van veranderingen in het karyotype met veranderingen in het fenotype., Mensen die bijvoorbeeld een extra chromosoom 21 hebben, hebben het syndroom van Down. Inserties, deleties en veranderingen in chromosoomaantal kunnen door de bekwame cytogeneticus worden gedetecteerd, maar het correleren van deze metspecifieke fenotypes is moeilijk.
de eerste onderscheidende parameter bij het ontwikkelen van een karyotype is de grootte en het aantal van de chromosomen. Hoewel dit nuttig is, biedt het niet voldoende detail om te beginnen met de ontwikkeling van een correlatie tussen structuur en functie (fenotype)., Om verder te onderscheiden tussen chromosomen, worden zij behandeld met een kleurstof die DNA op een reproduceerbare manier bevlekt. Nakleuring, sommige van de regio ‘ s zijn licht gekleurd en andere zijn zwaarbeschadigd. Zoals hierboven beschreven, zijn de licht gekleurde gebieden calledeuchromatin, en de donkere gekleurde regio heet heterochromatin. De huidige kleurstof van chose is de Giemsa vlek,en het resulterende patroon wordt het G-banding patroon genoemd.,
c-Waardeparadox
naast het beschrijven van het genoom van een organisme door het aantal chromosomen,wordt het ook beschreven door de hoeveelheid DNA in een haploïde cel. Dit wordt gewoonlijk uitgedrukt als de hoeveelheid DNA per haploïde cel (gewoonlijk uitgedrukt als picogrammen) of het aantal kilobases per haploïde cel en wordt de C-waarde genoemd. Oneimmediate eigenschap van eukaryotic organismen benadrukt een specifieke anomalie die vroeg in moleculair onderzoek werd ontdekt., Hoewel eukaryotic organismen 2-10 keer zo veel genen als prokaryotes lijken te hebben, hebben zij vele orde van grootte meer DNA in de cel. Bovendien is de hoeveelheid DNA per genoom gecorreleerd niet met de veronderstelde evolutionaire complexiteit van een soort.Dit wordt gesteld als de C-waardeparadox: de hoeveelheid DNA in de haploidcel van een organisme is niet gerelateerd aan zijn evolutionaire complexiteit. (Een ander belangrijk punt om in gedachten te houden is dat er geen relatie is tussen het aantal chromosomen en de veronderstelde evolutionaire complexiteit van een organisme.,)
C Values of Organisms Used in Genetic Studies
| Species | Kilobases/haploid genome |
|---|---|
| E. coli |
4.5 x 103 |
| Human |
3.0 x 106 |
| Drosophila |
1.7 x 105 |
| Maize |
2.0 x 106 |
| Aribidopsis |
7.,0 x 104 |
een dramatisch voorbeeld van het bereik van C-waarden kan worden gezien in het plantenrijk waar Arabidopsis de lage kant vertegenwoordigt en lelie (1,0 x 10^8 kb/haploidgenoom) de hoge kant van complexiteit. In gewicht uitgedrukt is dit 0,07 picogramsper haploïde Arabidopsis genoom en 100 picogrammen per haploïde lilygenoom.
genoom-de complete set chromosomen geërfd van een enkele ouder;de volledige DNA-component van een individu; de definitie sluit vaak organellen uit