Aitotumallisilla Kromosomin Rakenne
pituus DNA tumassa on paljon suurempi kuin koko compartmentin johon se sisältyy. Tähän lokeroon mahtuakseen DNA: n on täytynyt täyttyä jollain tavalla. DNA: n tiivistymisaste ilmaistaan sen pakkaussuhteessa.
Pakkaus suhde – pituus DNA jaettuna pituus whichit on pakattu
esimerkiksi, lyhin ihmisen kromosomi sisältää 4,6 x 107 bp DNA: ta(noin 10 kertaa genomin koko E. coli)., Tämä vastaa 14 000 µm: AA laajennettua DNA: ta. Mitoosin aikana kondensoituneimmassa tilassaan tekromosomi on noin 2 µm pitkä. Tällöin pakkaussuhde on 7 000(14 000/2).
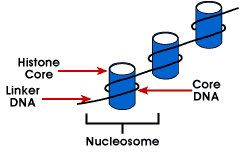
kokonaispakkaussuhteen saavuttamiseksi DNA: ta ei pakata suoraan kromatiinin finalstructureen. Sen sijaan se sisältää useita organisaatiohierarkioita.Ensimmäisen tason pakkaus on saavuttaa käämin DNA: n noin proteincore tuottaa ”helmi-kuin” – rakennetta kutsutaan nucleosome. Tämä luo pakkaussuhteen noin 6., Tämä rakenne on invariantti sekä kaikkien kromosomien teekromatiinissa että heterokromatiinissa. Toinen taso packingis coiling helmiä kierteiset rakenne nimeltään 30 nm fiberthat löytyy sekä interphase kromatiini ja kromosomit, mitoosi. Tämä rakenne lisää pakkaussuhdetta noin 40: een. Lopullinen pakkaus occurswhen kuitu on järjestetty silmukoita, tukirunkoja ja verkkotunnuksia, jotka antavat finalpacking suhde on noin 1000 interphase kromosomia ja noin 10 000 inmitotic kromosomit.,
Aitotumallisilla kromosomit koostuvat DNA-proteiini monimutkainen, joka on organizedin kompakti tavalla, joka mahdollistaa suuri määrä DNA: ta tulee säilyttää avaimia tuma solun. Kromosomin alayksikkö on kromatiini.Kromatiinin perusyksikkö on nukleosomi.,romatin – yksikön analyysin kromosomi; chromatin reflectsthe yleinen rakenne kromosomi, mutta ei ole ainutlaatuinen millään particularchromosome
Nucleosome – yksinkertaisin pakkaus DNA rakenne, joka on saatavana eukaryoottisesta kromosomeja; DNA on kääritty ympäri octamer pieniä basicproteins kutsutaan histones; 146 bp on kiedottu ydin ja remainingbases linkki seuraavaan nucleosome; tämä rakenne aiheuttaa negatiivisia supercoiling
nucleosome koostuu noin 200 bp kiedottu histoni octamerthat sisältää kaksi kappaletta histoni-proteiinien H2A, H2B, H3 ja H4., Nämä tunnetaan ydinhistorioina. Histonit ovat perusproteiineja, joilla on affinityfor DNA ja ovat runsaimpia DNA: han liittyviä proteiineja. Näiden neljän histonin aminoasidijakso säilyy, mikä viittaa samanlaiseen funktioon kaikille.
nukleosomiyksikköön liittyvän DNA: n pituus vaihtelee lajeittain. Koosta riippumatta kyse on kuitenkin kahdesta DNA-komponentista. CoreDNA on DNA, joka liittyy histonioktameeriin.Tämä arvo on invariantti ja on 146 emäsparia., Ydin-DNA muodostaa kaksi loopsaround, että octamer, ja tämä mahdollistaa kaksi aluetta, jotka ovat 80 bp toisistaan tobe tuodaan lähellä. Näin ollen kaksi kaukana toisistaan olevaa sekvenssiä voi toimia samalla säätelyproteiinilla kontrolloidakseen geenien ilmentymistä. TheDNA, joka on välillä kunkin Histoni oktameeri kutsutaan linker DNAand voi vaihdella pituus 8-114 emäsparit. Tämä vaihtelu on speciesspecific, mutta vaihtelu linkitys DNA: n pituus on myös liittynyt kanssa kehitysvaihe organismin tai tiettyjä alueita genomin.,
seuraavan tason organisaatio kromatiini on 30 nm kuitua. Tämä tarkoittaa solenoidirakennetta, jossa on noin 6 nukleosomia käännöstä kohti. Tällöin pakkaussuhde on 40, mikä tarkoittaa sitä, että jokainen 1 µm aksiskonferenssia pitkin saa 40 µm DNA: ta. Tämän rakenteen stabiilisuus edellyttää Histoni-geeniperheen viimeisen jäsenen, Histoni H1: n, esiintymistä., Becauseexperiments, että liuskan L1 alkaen chromatin säilyttää nucleosome, mutta ei 30 nm rakenne, pääteltiin, että H1 on tärkeää, että stabilizationof 30 nm rakenne.
pakkausten lopulliselle tasolle on ominaista 700 nm: n rakenne, joka näkyy metafaasi-kromosomissa. Tiivistetty pala kromatiini on acharacteristic rakennustelineet rakenne, joka voidaan havaita metaphasechromosomes. Tämä näyttää olevan seurausta dnain-kromosomin laajasta silmukoinnista.
viimeiset määritelmät, jotka on esitettävä, ovat eukromatiini ja meterokromatiini., Kun kromosomit värjätään väriaineilla, ne näyttävät muuttuvan kevyesti ja tummasti värjätyiksi alueiksi. Kevyesti värjätyt alueet ovat eukromatiinia ja sisältävät yhden kopion, geneettisesti aktiivista DNA: ta. Värjätyt alueet ovat heterokromatiinia ja sisältävät toistuvia sekvenssejä, jotka ovat geneettisesti inaktiivisia.
Sentromeerien ja Telomeerit
Sentromeerien ja telomeerit ovat kaksi tärkeää ominaisuuksia kaikkien eukaryoottisesta kromosomeja. Jokainen tarjoaa ainutlaatuisen funktion, joka on ehdottoman välttämätön kromosomin vakaudelle., Sentromeerien tarvitaan erottelua sentromeeri aikana meioosin ja mitoosin ja teleomeres tarjota terminaali vakautta kromosomi ja varmistaa sen säilymisen.
Sentromeerien ovat ne, tiivistetty alueiden kromosomi, jotka ovat vastuussa tarkan erottelun monistaa chromosomeduring mitoosin ja meioosin. Kun kromosomit värjätään, niissä näkyy tyypillisesti tumma alue, joka on centromere. Mitoosin aikana, sentromeeri, joka on jaettu sisar kromatidia täytyy jakaa niin, että kromatidia voi siirtää toopposite navat solu., Toisaalta, ensimmäisen meioottisen divisionthe sentromeeri sisar kromatidia on pysyttävä ehjänä, kun taas aikana meiosisII niiden on toimittava, koska ne eivät mitoosin aikana. Siksi sentromeeri on tärkeä osa kromosomin rakenteen ja erottelun.
Sisällä sentromeeri-alueen, useimmat lajit on useita paikkoja, joissa kara kuidut pitävät, ja nämä sivustot koostuvat DNA: ta sekä proteiinia. Varsinaista kiinnityskohtaa kutsutaan kinetokoreeksi ja se koostuu sekä DNA: sta että proteiinista. Näiden alueiden DNA-sekvenssiä kutsutaan DNA: ksi., Koska CEN DNA: ta voidaan siirtää yksi kromosomi toiseen ja silti kromosomi kyky erottaa nämä sekvenssit eivät saa tarjota minkään muun toiminnon.
Tyypillisesti CEN-DNA: ta on noin 120 emäsparin pitkä ja koostuu useista osa-alueet, CDE-en, CDE-II ja CDE-III. Mutaatiot kaksi ensimmäistä osa-alueet ei ole vaikutusta erottelun, mutta kohta mutaatio CDE-III osa-verkkotunnuksen poistaa kokonaan kyky sentromeeri aikana toimimaan kromosomi erottelu., Siksi CDE-III: n on osallistuttava aktiivisesti karan kuitujen sitomiseen sentromeeriin.
kinetochoren proteiinikomponenttia on vasta nyt luonnehdittu. Monimutkainen kolme proteiineja kutsutaan Cbf-III sitoutuu normaali CDE-III-alueilla, mutta ei voi sitoutua CDE-III-alueella, jossa on kohta mutaatio, joka estää mitoosi-erottelu. Lisäksi mutantit geenit koodaavat Cbf-III proteiineja myös poistaa kyky kromosomit erillään mitoosin aikana., Muita analyysejä sekä DNA: n ja proteiinin komponentteja sentromeeri on tarpeen täysin ymmärtää mekaniikka-kromosomi erottelu.
Telomeerit ovat DNA-alue, lopussa lineaarinen eukaryoticchromosome, että tarvitaan replikointi ja vakaus-kromosomi.McClintock tunnustettu niiden erityisiä ominaisuuksia, kun hän huomasi, että jos twochromosomes olivat rikki solu, pää voisi liittää toisaalta päinvastoin. Hän ei koskaan havainnut, että brokenend olisi kiinnittynyt katkeamattoman kromosomin päähän., Näin päät rikki chromosomesare tahmea, kun normaalisti loppuun ei ole tahmea, mikä viittaa siihen, päät ofchromosomes on ainutlaatuisia ominaisuuksia. Yleensä, mutta ei aina, telomeeridna on heterokromaattinen ja sisältää suoria tandemly toistuvia sekvenssejä. Seuraavassa taulukossa on esitetty useiden lajien toistosekvenssit. Nämä ovat usein muotoa (T/A)xGy, jossa x on välillä 1 ja 4 ja y on suurempi kuin 1.,
Telomere Repeat Sequences
| Species | Repeat Sequence |
|---|---|
| Arabidopsis | TTTAGGG |
| Human | TTAGGG |
| Oxytricha | TTTTGGGG |
| Slime Mold | TAGGG |
| Tetrahymena | TTGGGG |
| Trypanosome | TAGGG |
| Yeast | (TG)1-3TG2-3 |
Notice that the number of TG sequences and the number of cytosines in the yeast sequence varies., Ainakin hiiva, se on osoitettu, että eri kantoja sisältävät eri pituisia teleomeres ja että pituus on alle geneettistä valvontaa.
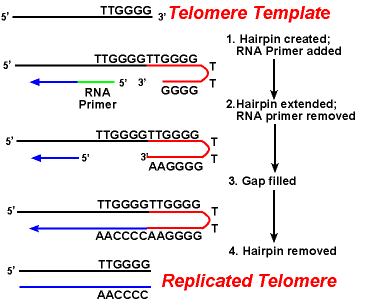
telomeerien ensisijainen vaikeus on lagging Strandin replikaatio.Koska DNA-synteesi vaatii RNA-malli (joka tarjoaa vapaa 3′-OHgroup) prime DNA: n replikaatio, ja tämä malli on lopulta hajonnut, lyhyt yksijuosteinen alueella olisi jäljellä lopussa kromosomi. Tämä alue olisi altis entsyymeille, jotka hajottavat yksijuosteista DNA: ta., Tuloksena olisi, että kromosomin pituus lyhenisi jokaisen jakovaiheen jälkeen. Mutta tätä ei nähdä.
toiminta telomeraasi-entsyymien varmistaa, että päät jäänyt osa on toistettu oikein. Hyvin tutkittu järjestelmä liittyy Tetrahymena alkueläin organismi. Tämän organismin telomeerit päättyvät jaksoon 5 ’- ttgggg-3’. Telomeraasi lisää joukon 5′-TTGGGG-3 ’ toistoja lagging Strandin päihin. Hiusneula syntyy, kun guaniinijäämien välillä esiintyy epätavallisia emäspareja toistuvassa muodossa., Seuraavaksi RNA-pohjamaali poistetaan, ja lagging-juosteen 5 ’ päätä voidaan käyttää DNA-synteesiin. Ligaatio tapahtuu valmiin lagging Strandin ja hiusneulan välillä. Lopuksi hiusneula poistetaan kohdassa 5′-TTGGGG-3’ Toista. Näin kromosomin loppu toistuu uskollisesti. Seuraavassa kuvassa esitetään nämä vaiheet.,
Replikointi Telomeerit
Analyysi DNA-Sekvenssit Eukaryoottisesta Genomin
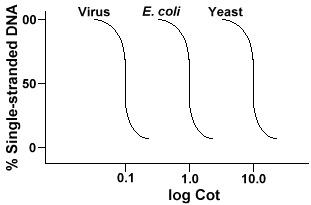
tekniikka, jota käytetään määrittää järjestyksen monimutkaisuus tahansa genomeinvolves, että denaturointi ja renaturation DNA: ta. DNA on denaturedby-lämmitys, joka sulattaa h-sidokset ja muuttaa DNA: n yksijuosteiseksi. Jos DNA jäähtyy nopeasti, DNA jää yksijuosteiseksi. Mutta jos DNAis antaa jäähtyä hitaasti, toisiaan täydentävät sekvenssit löytävät toisensa ja lopulta emäsparin uudelleen., DNA: n uudelleensijoittumisnopeus(toinen nimitys renatuurille) on sen lajin funktio, josta Dnawat eristäytyivät. Alla on käyrä, joka saadaan yksinkertaisesta genomista.
Y-akseli on se prosentti DNA: sta, joka jää yksijuosteiseksi. Tämä isexpressed suhteena pitoisuus yksijuosteisen DNA: n (C)yhteensä pitoisuus alkaa DNA (Co). X-akselin isa-log-asteikolla tuotteen alkuperäinen pitoisuus DNA: n (mol/l)saadaan kertomalla pituus kertaa reaktio eteni (sekunneissa)., Tämän arvon määrittely on Cot ja sitä kutsutaan ” Cot ” – arvoksi. Käyrää itseään kutsutaan ” Cot ” – käyräksi. Kuten voidaan nähdä, käyrä on melko sileä, mikä osoittaa, että uudelleenjulkistaminen tapahtuu hidastuu, mutta vähitellen ajan kuluessa. Yksi hyödyllinen erityisarvo on cot½, jossa puolet DNA: sta on uudelleenjulkaistu.
DNA: n denaturaatio-ja Renaturaatiokokeisiin liittyvät vaiheet
1. Perimä on noin 400 bp.
2. Denaturoi DNA kuumentamalla 100oC: ksi.
3. Jäähdytä hitaasti ja ota näytteitä eri aikavälein.
4., Määritetään yksijuosteisen DNA: n osuus kussakin vaiheessa.
muoto ”Pinnasänky” käyrä tiettyä lajia varten on tehtävä kaksi tekijää:
- koko tai monimutkaisuus genomin; ja
- määrä toistuvia DNA: n sisällä genomin
Jos me piirtää ”Pinnasänky” käyrät genomin kolme lajia, kuten bacteriophagelambda, E. coli ja hiiva näemme, että niillä on sama muoto,mutta Cot½ hiiva on suurin, E. coli nextand lambda pienin., Fyysisesti, mitä suurempi genomin koko, sitä kauemmin se kestää yhden sekvenssin kohdata sen täydentävä sekvenssi ratkaisun. Tämä johtuu siitä, että kahden toisiaan täydentävän sekvenssin on kohdattava toisensa ennen kuin ne voivat pariutua. Monimutkaisempi genomin, joka on themore ainutlaatuisia sekvenssejä, jotka ovat käytettävissä, sitä kauemmin se kestää anytwo täydentäviä sekvenssit kohtaavat toisensa ja pari. Kun otetaan huomioon vastaavanlaiset ratkaisut, Cot½: n saavuttamiseen tarvitaan monimutkaisempia lajeja.,
Toistuvat DNA-sekvenssit, DNA-sekvenssit, jotka löytyvät useammin kuin kerran inthe genomin laji, on erottuva vaikutuksia ”Pinnasänky” käyrät.Jos tietty sekvenssi on edustettuna kaksi kertaa genomin se on twocomplementary sekvenssit parin kanssa ja siten on Pinnasänky arvo kaksi kertaa vähemmän suuri kuin sekvenssi edustaa vain kerran genomin.
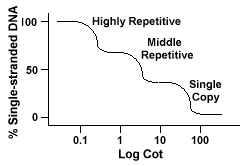
Eukaryoottisilla genomeilla on todellisuudessa laaja joukko sekvenssejä, jotka edustavat eri toistotasoja., Yhden kopion sekvenssit löytyvät onceor muutaman kerran perimässä. Monet sekvenssit, jotka koodaavat funktionalgenes kuuluvat tähän luokkaan. Keskimmäistä toistuvaa DNA: ta löytyy 10s -1000 kertaa perimästä. Esimerkkejä näistä olisivat rRNA ja trnageenit sekä varastointiproteiinit kasveissa, kuten maississa. Lähi toistuvia DNAcan vaihtelevat 100-300 bp 5000 bp, ja se voi olla hajallaan koko genomin.Runsaimmat sekvenssit löytyvät erittäin toistuvasta Dnaclassista., Näitä sekvenssejä esiintyy genomeandissa 100 000-1 miljoonaa kertaa, ja ne voivat vaihdella kooltaan muutamasta useisiin satoihin emästen pituuteen. Thesesequences löytyy alueiden kromosomi kuten heterochromatin,sentromeerien ja telomeerit ja ovat yleensä järjestetty tandem toistaa. Seuraavassa on esimerkki tandemly toistuva sekvenssi:
ATTATA ATTATA ATTATA // ATTATA
Genomit, jotka sisältävät nämä eri luokat sekvenssit reanneal vuonna differentmanner kuin genomien kanssa vain yhden kopion sekvenssit., Sen sijaan yksi sileä ”Pinnasänky” – käyrä, kolme eri käyrät voidaan nähdä, kukin representinga eri toisto-luokan. Ensimmäiset sekvenssit uudelleenjulkistamiseen ovat korkearepetitiiviset sekvenssit, koska niistä on niin paljon kopioita genomissa ja koska niillä on alhainen sekvenssin monimutkaisuus. Toinen osa thegenomesta uudelleenjulkaistavaksi on keskimmäinen toistuva DNA, ja viimeinen osa toreanneal on yhden kopion DNA., Seuraava kaavio kuvaa ”Pinnasänky”käyrä ”tyypillinen” eukaryoottisesta genomin
seuraavassa taulukossa on esitetty järjestyksessä jakelu valittu lajeja.
| Lajit | – Sekvenssi Jakelu |
|---|---|
| Bakteerit | 99.,27% Lähi-Toistuvia 8% Erittäin Toistuvia |
| Maissi | 30% Yhden Kopioida 40% Lähi Toistuvia 20% Erittäin Toistuvia |
| Vehnä | 10% Yhden Kopioida 83% Lähi Toistuvia 4% Erittäin Toistuvia |
| Arabidopsis | 55% irtonumeromyynti 27% Lähi-Toistuvia 10% Erittäin Toistuvia |
Sarja Interspersion
vaikka genomien eliöt sisältävät yhden kopion, middlerepetitive ja erittäin toistuvia DNA-sekvenssejä, nämä sekvenssit eivät ole arrangedsimilarly kaikki lajit., Näkyvää järjestelyä kutsutaan lyhyeksi periodintersioniksi. Tämä järjestely on ominaista toistuva sequences100-200 bp pituus välissä keskuudessa yhtenä kappaleena sekvenssit, jotka ovat 1000-2000bp pituus. Tätä järjestelyä esiintyy eläimissä, sienissä ja kasveissa.
toinen asetelmatyyppi on pitkän ajan interspersio. Tämä on ominaista 5000 bp ulottuu toistuvat sekvenssit interspersedwithin alueet 35 000 bp yhden kopioida DNA: ta. Drosophila on lajitutkintolaji, jolla on tämä melko harvinainen sekvenssijärjestely., Molemmissa tapauksissa, therepeated sekvenssit ovat yleensä keski toistuva Luokka. Keskustelimme siitä, missä esiintyy hyvin toistuvia sekvenssejä.
Aitotumallisilla Kromosomi Karyotyyppi
ottaa huomioon, Että bakteereja on vain yksi kromosomi, eukaryoottisten lajien on ainakin yhdet kromosomeja. Useimmilla on useampi kuin yksi pari. Toinen relevantti seikka on, että eukaryoottisia kromosomeja havaitaan vain solujen jakautumisen aikana eikä solusyklin kaikissa vaiheissa. Ne ovat metafaasin aikana pahimmillaan sisarkromatidien kiinnittyessä.,Tämä on sytogeneettisen analyysin ensisijainen vaihe.
jokaiselle lajille on ominaista karyotyyppi. Karyotyyppi on adescription määrä kromosomeja normaalissa diploidi solu, sekä niiden kokojakauma. Esimerkiksi ihmisen kromosomissa on 23 pairsof-kromosomia, 22 somaattista paria ja yksi sukupuolikromosomipari. Yksi tärkeä geenitutkimuksen osa on karyotyypin muutosten korreloiminen yksilön fenotyypin muutoksiin.
yksi genetiikan tärkeä osa-alue korreloi karyotyypin muutosten kanssa fenotyypissä., Esimerkiksi ihmisillä, joilla on ylimääräinen kromosomi 21have Downin oireyhtymä. Lisäykset, poistot ja muutokset kromosomi numbercan havaita taitava cytogeneticist, mutta korreloivat joissaerityinen fenotyyppeihin on vaikeaa.
ensimmäinen erotteleva parametri karyotyypin kehittyessä on kromosomien koko ja lukumäärä. Vaikka tämä on hyödyllinen, se ei provideenough yksityiskohtia voidaan alkaa kehittää korrelaatio rakenneja toiminta (fenotyyppi)., Jotta kromosomit voitaisiin edelleen erottaa toisistaan, niitä käsitellään väriaineella, joka tahraa DNA: n toistettavalla tavalla. Jälkipuinti, jotkut alueet ovat kevyesti värjätty ja toiset ovat raskaasti. Kuten edellä on kuvattu, kevyesti värjätään alueet ovat calledeuchromatin, ja tumma petsattu alue on calledheterochromatin. Nykyinen väriaine on Giemsa tahra, ja tuloksena kuvio kutsutaan G-ryhmittely kuvio.,
C-Arvon Paradoksi.
lisäksi kuvataan genomin organismin sen määrä kromosomeja,se on myös kuvattu määrä DNA haploidi solu. Tämä on usuallyexpressed määrä DNA kohti haploidi solu (yleensä ilmaistuna pg)tai määrä kilobases per haploidi solu ja on nimeltään C-arvo. Eukaryoottisten eliöiden pikaominaisuus korostaa erityistä poikkeamaa, joka havaittiin molekyylitutkimuksen varhaisessa vaiheessa., Vaikka eukaryoottisilla järjestöillä on 2-10 kertaa niin paljon geenejä kuin prokaryooteilla, niillä on paljon suuruusluokkaa enemmän DNA: ta solussa. Lisäksi DNA: n määrä genomeita kohti ei korreloi lajin oletetun evolutiivisen monimutkaisuuden kanssa.Tämä on todettu C-arvon paradoksi: määrä DNA: ta haploidcell organismin ei liity sen evoluution monimutkaisuus. (Toinen tärkeä seikka pitää mielessä on, että ei ole suhdetta välinen kromosomien lukumäärän ja oletetun evoluution monimutkaisuus anorganism.,)
C Values of Organisms Used in Genetic Studies
| Species | Kilobases/haploid genome |
|---|---|
| E. coli |
4.5 x 103 |
| Human |
3.0 x 106 |
| Drosophila |
1.7 x 105 |
| Maize |
2.0 x 106 |
| Aribidopsis |
7.,0 x 104 |
dramaattinen esimerkki valikoima C arvot voidaan nähdä kasvi kingdomwhere Arabidopsis edustaa low end ja lily (1,0 x 10^8 kb/haploidgenome) high-end monimutkaisuus. Painon kannalta tämä on 0.07 picogramsper haploidi Arabidopsis genomin ja 100 pg per haploidi lilygenome.
Genome – täydellinen joukko kromosomit periytyvät yksinhuoltaja;täydellinen DNA-osa yksilön; määritelmä usein excludesorganelles