Eukaryote Kromosom Struktur
lengden av DNA i kjernen er langt større enn størrelsen på compartmentin der det finnes. For å passe inn i dette rommet DNA har å becondensed på noen måte. I hvilken grad DNA er kondensert er expressedas sin pakking forhold.
Pakking ratio – lengden av DNA dividert med lengden i whichit er pakket
For eksempel, den korteste menneskelig kromosom inneholder 4,6 x 107 bp DNA(ca 10 ganger genomstørrelse av E. coli)., Dette er tilsvarende til 14,000 µm av utvidet DNA. I sin mest komprimert tilstand ved mitose, thechromosome er om 2 µm lange. Dette gir et pakking forholdet mellom 7000(14,000/2).
for Å oppnå den samlede pakking forhold, DNA er ikke pakket direkte inn finalstructure av chromatin. I stedet, det inneholder flere hierarkier av organisasjonen.Det første nivået av pakking oppnås ved avvikling av DNA rundt en proteincore til å produsere en «perle» lignende struktur som kalles en nucleosome. Thisgives pakking et forhold på omtrent 6., Denne strukturen er invariant i både theeuchromatin og heterochromatin av alle kromosomer. Det andre nivået av packingis den coiling av perler i en spiralformet struktur kalt 30 nm fiberthat er funnet i både interphase chromatin og mitotic kromosomer. Thisstructure øker pakking forholdet til ca 40. Den endelige emballasje occurswhen fiber er organisert i looper, stillaser og domener som gir en finalpacking ratio på ca 1000 i interphase kromosomer, og ca 10 000 inmitotic kromosomer.,
Eukaryote kromosomene består av et DNA-protein kompleks som er organizedin en kompakt måte som tillater den store mengden av DNA for å bli lagret i kjernen av cellen. Den subunit betegnelse på kromosom er chromatin.Den grunnleggende enhet chromatin er nucleosome.,romatin – enhet for analyse av kromosom; chromatin reflectsthe generelle strukturen av kromosom, men er ikke unike for alle particularchromosome
Nucleosome – enkleste emballasje strukturen av DNA som er funnet inall eukaryote kromosomer; DNA er pakket rundt en octamer av små basicproteins kalt histones; 146 bp er pakket rundt kjernen og remainingbases link til neste nucleosome; denne strukturen fører til negative supercoiling
nucleosome består av om lag 200 bp pakket rundt en histone octamerthat inneholder to kopier av histone proteiner H2A, H2B, H3 og H4., Disse areknown som kjernen histones. Histones er grunnleggende proteiner som har en affinityfor DNA og er den mest tallrike protein som er forbundet med DNA. Den aminoacid sekvens av disse fire histones er bevart tyder på en lignende functionfor alle.
lengden av DNA som er forbundet med nucleosome enhet varierer betweenspecies. Men uavhengig av størrelsen, to DNA-komponenter er involvert. CoreDNA er DNA som er faktisk forbundet med histone octamer.Denne verdien er invariant og er 146 base-par., Kjerne-DNA danner to loopsaround den octamer, og dette tillater at to regioner som er 80 bp bortsett tobe brakt inn i umiddelbar nærhet. Dermed, to sekvenser som er langt fra hverandre caninteract med de samme regulatoriske proteiner for å kontrollere genuttrykk. TheDNA som er mellom hver histone octamer kalles linker DNAand kan variere i lengde, fra 8 til 114 base-par. Denne varianten er speciesspecific, men variasjon i linker DNA-lengde har også vært forbundet med utviklingsmessige fasen av organismen eller bestemte regioner av genomet.,
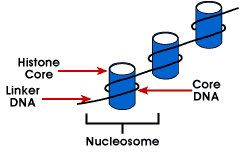
neste nivå av organisering av chromatin er 30 nm fiber. Thisappears å være en solenoid struktur med ca 6 nucleosomes per tur. Thisgives et pakking forholdet mellom 40, noe som betyr at hver 1 µm langs axiscontains 40 µm av DNA. Stabiliteten av denne strukturen krever thepresence av den siste medlem av histone genet familie, histone H1., Becauseexperiments at stripen H1 fra chromatin opprettholde nucleosome, men notthe 30 nm struktur, og det ble konkludert med at H1 er viktig for stabilizationof 30 nm struktur.
Det endelige nivået av emballasje er preget av 700 nm struktur seenin den metaphase kromosom. Sammendratt stykke chromatin har acharacteristic stillas struktur som kan bli oppdaget i metaphasechromosomes. Dette ser ut til å være et resultat av omfattende looping av DNAin kromosomet.
Den siste definisjoner som trenger å bli presentert er euchromatin andheterochromatin., Når kromosomer er farget med fargestoffer, de ser ut til å havealternating lett og mørk farget regioner. Lett-farget regionsare euchromatin og inneholde enkelt-kopi, genetisk-aktiv DNA. Thedarkly-farget regioner er heterochromatin og inneholder repeterende sequencesthat er genetisk inaktive.
Centromeres og Telomeres
Centromeres og telomeres er to viktige funksjoner i alle eukaryote kromosomer. Hver tilbyr en unik funksjon som er helt nødvendig for stabiliteten i kromosomet., Centromeres er nødvendig for segregering av centromere under meiose og mitose, og teleomeres gi terminal stabilitet til kromosom og sikre sin overlevelse.
Centromeres er de kondensert regioner i kromosom thatare ansvarlig for nøyaktige segregering av replikert chromosomeduring mitose og meiose. Når kromosomer er beiset de vanligvis viser en mørk-farget område som er centromere. Under mitosen, den centromere som deles av søster kromatidene må dele slik at kromatidene kan migrere toopposite polene i cellen., På den annen side, i løpet av de første meiotic divisionthe centromere av søster kromatidene må forbli intakt, mens under meiosisII de må handle som de gjør i løpet av cellesyklus. Derfor centromere er animportant del av kromosom struktur og segregering.
Innenfor centromere regionen, de fleste arter har flere steder hvor spindel fibre fest, og disse områdene består av DNA samt proteiner. Den faktiske plasseringen der vedlegget oppstår kalles kinetochore og er sammensatt av både DNA og protein. DNA-sekvensen i disse regionene er calledCEN DNA., Fordi CEN DNA kan flyttes fra et kromosom til et annet og fortsatt gi kromosom med evnen til å skille, disse sekvensene må ikke gi noen annen funksjon.
Vanligvis CEN DNA er ca 120 base par lang og består av flere sub-domener, CDE-jeg, CDE-II og CDE-III. Mutasjoner i de to første sub-domener har ingen effekt på segregering, men et punkt mutasjon i CDE-III sub-domene helt eliminerer muligheten for centromere til å fungere i løpet av kromosom segregering., Derfor CDE-III må være aktivt involvert i binding av spindel fiber til centromere.
protein del av kinetochore er først nå blir preget. Et kompleks av tre proteiner som kalles Cbf-III binder seg til normal CDE-III-regioner, men kan ikke binde seg til en CDE-III-regionen med et punkt mutasjon som hindrer mitotic segregering. Videre, mutanter av gener koding av Cbf-III proteiner fjerner også muligheten for kromosomer å skille under mitosen., Ytterligere analyser av DNA og protein komponenter av centromere er nødvendig for å fullt ut forstå mekanikken av kromosom segregering.
Telomeres er den regionen av DNA på slutten av lineær eukaryoticchromosome som er nødvendig for replikasjon og stabilitet av kromosomet.McClintock anerkjent sine spesielle funksjoner når hun la merke til at hvis twochromosomes ble ødelagt i en celle, slutten av en kunne legge til otherand vice versa. Hva hun har aldri observert var feste av brokenend til slutten av en ubrutt kromosom., Dermed ender brutt chromosomesare klissete, mens normal slutten er ikke klebrig, noe som tyder på endene ofchromosomes har unike funksjoner. Vanligvis, men ikke alltid, telomericDNA er heterochromatic og inneholder direkte tandemly repeterte sekvenser. Thefollowing tabellen viser gjenta sekvensen av flere arter. Disse er ofte av skjemaet (T/A)xGy der x er mellom 1 og 4 og y er større enn 1.,
Telomere Repeat Sequences
| Species | Repeat Sequence |
|---|---|
| Arabidopsis | TTTAGGG |
| Human | TTAGGG |
| Oxytricha | TTTTGGGG |
| Slime Mold | TAGGG |
| Tetrahymena | TTGGGG |
| Trypanosome | TAGGG |
| Yeast | (TG)1-3TG2-3 |
Notice that the number of TG sequences and the number of cytosines in the yeast sequence varies., I hvert fall for gjær, det har vist seg at ulike stammer inneholder ulike lengder av teleomeres og at lengden er under genetisk kontroll.
Den primære problemer med telomeres replikering av lagging strand.Fordi DNA-syntese krever en RNA-mal (som gir den fri 3′-OHgroup) prime DNA replikasjon, og denne malen er slutt degradert, en kort single-strandet regionen ville bli igjen på slutten av kromosomene. Dette området vil være utsatt for enzymer som bryter ned enkelt-strandet DNA., Resultatet vil være at lengden på kromosom ville bli forkortet etter hver divisjon. Men dette er ikke synlig.
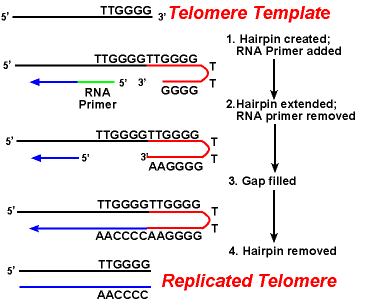
virkningen av telomerase enzymer sikre at endene av henger trådene er replikert på riktig måte. En godt studert systemet innebærer Tetrahymena protozoer organismen. Telomeres av denne organismen i slutten av sekvensen 5′-TTGGGG-3′. Den telomerase legger til en serie av 5′-TTGGGG-3′ gjentar til endene av lagging strand. En hårnål oppstår når uvanlige base-par mellom guanin rester i gjenta form., Neste RNA-primer er fjernet, og 5′ – enden av lagging strand kan brukes for DNA-syntese. Ligation oppstår mellom de ferdig lagging strand og skarpe. Til slutt, hårnål er fjernet ved 5′-TTGGGG-3′ gjenta. Dermed enden av kromosomet er trofast replikert. Følgende figur viser disse trinnene.,
Replikering av Telomeres
Analyse av DNA-Sekvenser i Eukaryote Genomer
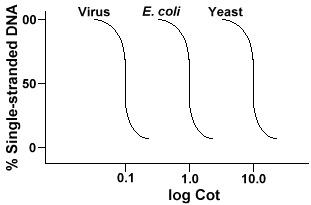
Den teknikken som er brukt for å bestemme rekkefølgen kompleksiteten av noen genomeinvolves den denaturering og renaturation av DNA. DNA er denaturedby oppvarming som smelter H-obligasjoner og gjør DNA-single-strandet. Ifthe DNA blir raskt nedkjølt, DNA forblir single-strandet. Men hvis DNAis kjøle seg sakte, sekvenser som er komplementære, vil finne hverandre og til slutt base par igjen., Den hastigheten som de DNA reanneals(et annet begrep for renature) er en funksjon av arter som DNAwas isolert. Nedenfor er en kurve som er innhentet fra en enkel genom.
Y-aksen er prosent av DNA-et som er enkelt strandet. Dette isexpressed som et forhold mellom konsentrasjonen av enkelt-strandet DNA (C)til den totale konsentrasjonen av start-DNA (Co). X-aksen isa log-skala av produktet til den opprinnelige konsentrasjonen av DNA (i føflekker/liter)multiplisert med lengden av tid reaksjonen forløp (i sekunder)., Den designationfor denne verdien er Barneseng og er kalt «Vugge» verdi. Kurven itselfis kalt en «Vugge» kurve. Som kan sees kurven er ganske glatt whichindicates at reannealing oppstår bremse, men gradvis over en periode oftime. En bestemt verdi som er nyttig er Cot½ , theCot verdi, hvor halvparten av DNA har reannealed.
Trinnene som er Involvert i DNA-Denaturering og Renaturation Eksperimenter
1. Skjær DNA til en størrelse på ca 400 bp.
2. Denaturering av DNA ved oppvarming til 100oC.
3. Sakte kule og ta prøver på forskjellige tidsintervaller.
4., Avgjøre % single-etterlatt DNA på hvert tidspunkt.
form av en «Vugge» kurve for en gitt art er en funksjon av to faktorer:
- størrelse eller kompleksitet av genom, og
- mengden av repeterende DNA i genomet
Hvis vi plotte «Vugge» kurver av genom av tre arter som bacteriophagelambda, E. coli og gjær vi vil se at de har samme form,men Cot½ av gjær vil bli størst, E. coli nextand lambda minste., Fysisk, jo større genomstørrelse lengre itwill ta for en hvilken som helst rekkefølge for å møte sin komplementære sekvens inthe løsning. Dette er fordi to komplementære sekvenser må encountereach andre før de kan koble sammen. De mer komplekse genom, som er themore unike sekvenser som er tilgjengelig, jo lenger tid vil det ta for anytwo komplementære sekvenser til å møte hverandre og par. Gitt similarconcentrations i løsningen, vil det da ta en mer komplekse arter longerto nå Cot½ .,
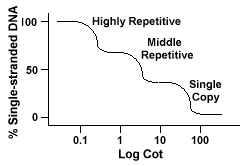
Gjentatte DNA-sekvenser, DNA-sekvenser som er funnet mer enn en gang i genomet av arten har karakteristiske virkninger på «Sengen» kurver.Hvis det er en bestemt sekvens er representert to ganger i genom vil det ha twocomplementary sekvenser hvis du vil sammenkoble med, og som sådan vil ha en Barneseng verdi halfas stor som en sekvens representerte bare en gang i genomet.
Eukaryote genomer faktisk har et bredt utvalg av sekvenser som er representedat ulike nivåer av repetisjon., Enkelt kopi sekvenser er funnet onceor et par ganger i genomet. Mange av sekvenser som koder for functionalgenes faller inn i denne klassen. Midt repeterende DNA er funnet fra 10s og 1000 ganger i genomet. Eksempler på disse vil omfatte rRNA og tRNAgenes og lagring av proteiner i planter som mais. Midt repeterende DNAcan variere fra 100-300 bp til 5000 bp og kan være spredt over hele genomet.De mest tallrike sekvenser er funnet i svært repeterende DNAclass., Disse sekvensene er funnet 100 000 til 1 million ganger i genomeand kan variere i størrelse fra noen få til flere hundre baser i lengden. Thesesequences er funnet i områder av kromosomet som heterochromatin,centromeres og telomeres og har en tendens til å bli arrangert som en tandem gjentar. Thefollowing er et eksempel på en tandemly gjentatt rekkefølge:
ATTATA ATTATA ATTATA // ATTATA
Genomet som inneholder disse forskjellige klasser av sekvenser reanneal i en differentmanner enn genomer med bare én kopi sekvenser., I stedet for å ha asingle glatt «Vugge» kurve, tre forskjellige kurver kan bli sett, hver representinga forskjellige repetisjon klasse. Den første sekvenser for å reanneal er highlyrepetitive sekvenser fordi så mange kopier av dem eksisterer i genom,og fordi de har en lav sekvens kompleksitet. Den andre delen av thegenome å reanneal er midt repeterende DNA, og den siste delen toreanneal er enkelt kopi av DNA-et., Følgende diagram viser de «Vugge»kurve for en «typisk» eukaryote genom
følgende tabell gir sekvensen distribusjon for utvalgte arter.
| Arter | Sekvens Distribusjon |
|---|---|
| Bakterier | 99.,27% Midten Repeterende 8% Svært Repeterende |
| Mais | 30% Enkelt Kopi 40% Midten Repeterende 20% Svært Repeterende |
| Hvete | 10% Enkelt Kopi 83% Midten Repeterende 4% Svært Repeterende |
| Arabidopsis | 55% Enkelt Kopi 27% Midten Repeterende 10% Svært Repeterende |
Sekvens Interspersion
Selv om genomet til høyere organismer inneholde ett eksemplar, middlerepetitive og svært repeterende DNA-sekvenser, disse sekvensene er ikke arrangedsimilarly i alle arter., Den fremtredende ordningen kalles kort periodinterspersion. Denne ordningen er preget av gjentatte sequences100-200 bp i lengde ispedd blant enkelt kopi av sekvenser som er 1000-2000bp i lengden. Denne ordningen er funnet i dyr, sopp og planter.
Den andre type arrangement er lang periode interspersion. Thisis preget av 5000 bp strekninger av repeterte sekvenser interspersedwithin regioner på 35 000 bp av én kopi av DNA-et. Drosophila er en exampleof en art med dette uvanlig sekvens arrangement., I begge tilfeller, therepeated sekvenser er vanligvis fra midten repeterende klasse. Vi discussedabove hvor svært repeterende sekvenser er funnet.
Eukaryote Kromosom Karyotype
Mens bakterier har bare en enkelt kromosom, eukaryote arter har minst ett par av kromosomer. De fleste har mer enn ett par. En annen relevantpoint er at eukaryote kromosomer er oppdaget bare skje under celldivision og ikke i alle faser av celle syklus. De er i sin mostcondensed skjemaet under metaphase når søsteren kromatidene er vedlagt.,Dette er den primære stadium når cytogenetic analyse er utført.
Hver art er preget av en karyotype. Den karyotype er adescription av antall kromosomer i normal diploid celle, som wellas deres størrelse distribusjon. For eksempel, menneskelig kromosom har 23 pairsof kromosom, 22 somatiske par og ett par av sex-kromosomer. En importantaspect av genetisk forskning er å sammenholde endringer i karyotype med changesin phenotype av den enkelte.
En viktig del av genetikk er å sammenholde endringer i karyotype withchanges i fenotypen., For eksempel, mennesker som har et ekstra kromosom 21have Down syndrom. Innsettinger, slettinger og endringer i kromosom numbercan bli oppdaget av dyktige cytogeneticist, men å sammenholde disse withspecific phenotypes er vanskelig.
Den første kritiske parameter når du utvikler en karyotype er sizeand antall kromosomer. Selv om dette er nyttig, er det ikke provideenough detalj for å begynne utviklingen av en korrelasjon mellom structureand funksjon (fenotypen)., For ytterligere å skille mellom kromosomer, theyare behandlet med et fargestoff som flekker DNA i en reproduserbar måte. Afterstaining, noen av regionene er lett farget og andre er heavilystained. Som beskrevet ovenfor, lett farget regioner er calledeuchromatin, og mørk farget regionen er calledheterochromatin. Gjeldende fargestoff av valgte er Giemsa flekken,og den resulterende mønsteret kalles G-mønster.,
C-Verdi Paradoks
I tillegg til å beskrive genom av en organismen med dens antall kromosomer,det er også beskrevet av mengden av DNA i en haploid celle. Dette er usuallyexpressed som mengden av DNA-per haploid celle (vanligvis uttrykt som picograms)eller antall kilobases per haploid celle og kalles C-verdi. Oneimmediate funksjon av eukaryote organismer fremhever en bestemt anomali thatwas oppdaget tidlig i molekylær forskning., Selv om eukaryote organismsappear å ha 2-10 ganger så mange gener som prokaryotes, de har mange ordersof størrelsesorden mer DNA i cellen. Videre er mengden av DNA per genomeis korrelerte ikke med den antatte evolusjonære kompleksiteten av en art.Dette er oppgitt som C-verdi paradoks: mengden av DNA i haploidcell av en organisme er ikke knyttet til dens evolusjonære kompleksitet. (Anotherimportant poeng å huske på er at det er ingen sammenheng betweenthe antall kromosomer og den antatte evolusjonære kompleksiteten av anorganism.,)
C Values of Organisms Used in Genetic Studies
| Species | Kilobases/haploid genome |
|---|---|
| E. coli |
4.5 x 103 |
| Human |
3.0 x 106 |
| Drosophila |
1.7 x 105 |
| Maize |
2.0 x 106 |
| Aribidopsis |
7.,0 x 104 |
En dramatisk eksempel på omfanget av C-verdier kan sees i anlegget kingdomwhere Arabidopsis representerer den lave enden og lily (1,0 x 10^8 kb/haploidgenome) den høye enden av kompleksitet. I vekt vilkårene dette er 0.07 picogramsper haploid Arabidopsis genom og 100 picograms per haploid lilygenome.
Genom – komplett sett av kromosomer nedarves fra en forelder, den komplette DNA-komponent til et individ; definisjonen ofte excludesorganelles