eukaryotisk kromosomstruktur
længden af DNA i kernen er langt større end størrelsen af det kompartment, hvori det er indeholdt. For at passe ind i dette rum skal DNA ‘ et kondenseres på en eller anden måde. Graden af hvilken DNA er kondenseret udtrykkessom dets pakkeforhold.
Pakkeforhold – længden af DNA divideret med længden i hvilkenDen er pakket
for eksempel indeholder det korteste humane kromosom 4,6 107 107 bp DNA(ca.10 gange Genomstørrelsen af E. coli)., Dette svarer til 14.000 µm udvidet DNA. I sin mest kondenserede tilstand under mitose, denkromosom er omkring 2 µm lang. Dette giver et pakkeforhold på 7000 (14.000/2).
for at opnå det samlede pakkeforhold pakkes DNA ikke direkte ind i finalestructure of chromatin. I stedet indeholder den flere hierarkier af organisation.Det første niveau af pakning opnås ved vikling af DNA omkring en proteincore for at producere en “perlelignende” struktur kaldet et nukleosom. Dette giver et pakkeforhold på omkring 6., Denne struktur er invariant i både deneuchromatin og heterochromatin af alle kromosomer. Det andet niveau af pakning er spolen af perler i en spiralformet struktur kaldet 30 nm fiberdet findes i både interfase chromatin og mitotiske kromosomer. Dettestruktur øger pakkeforholdet til omkring 40. Den endelige emballage occurswhen fiber er organiseret i loops, stilladser og domæner, som giver en finalpacking ratio på omkring 1000 i interphase kromosomer og omkring 10.000 inmitotic kromosomer.,
eukaryote kromosomer består af et DNA-proteinkompleks, der er organiseret på en kompakt måde, der tillader, at den store mængde DNA opbevares i cellekernen. Underenhedsbetegnelsen for kromosomet er kromatin.Den grundlæggende enhed af kromatin er nukleosomet.,romatin – enhed for analyse af kromosom; kromatin reflectsthe generelle struktur af kromosom, men er ikke unik for enhver particularchromosome
Nucleosome – enkleste emballage struktur af DNA, der er fundet inall eukaryote kromosomer; DNA, der er pakket omkring en octamer af små basicproteins kaldet histoner; 146 bp er viklet rundt om kernen og remainingbases link til det næste nucleosome; denne struktur medfører negative supercoiling
nucleosome består af omkring 200 bp viklet omkring en histon octamerthat indeholder to kopier af histon proteiner, H2A, H2B, H3 og H4., Disse er kendt som de centrale histoner. Histoner er basale proteiner, der har en affinitetfor DNA og er de mest rigelige proteiner forbundet med DNA. Aminosyresekvensen af disse fire histoner bevares, hvilket antyder en lignende funktion for alle.
længden af DNA, der er forbundet med nukleosomenheden, varierer mellemarter. Men uanset størrelsen er to DNA-komponenter involveret. CoreDNA er det DNA, der faktisk er forbundet med histonoctamer.Denne værdi er invariant og er 146 basepar., Kernen DNA danner to sløjferomkring octamer, og dette tillader to regioner, der er 80 bp fra hinanden tobe bragt i umiddelbar nærhed. Således kan to sekvenser, der er langt fra hinanden, interagere med det samme regulatoriske protein for at kontrollere genekspression. Dendna, der er mellem hver histonoktamer kaldes linker Dnaog kan variere i længde fra 8 til 114 basepar. Denne variation er speciesspecifik, men variation i linker DNA længde har også været forbundet medudviklingsstadiet af organismen eller specifikke regioner af genomet.,
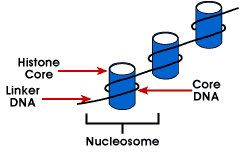
Det næste niveau af organisation af kromatin er 30 nm fibre. Dettevises at være en solenoidstruktur med omkring 6 nukleosomer pr. Dettegiver et pakningsforhold på 40, hvilket betyder, at hver 1 µm langs aksenindeholder 40 µm DNA. Stabiliteten af denne struktur kræverTilstedeværelsen af det sidste medlem af histongenfamilien, histon H1., Fordieksperimenter, der striber H1 fra kromatin, opretholder nukleosomet, men ikke 30 nm strukturen, blev det konkluderet, at H1 er vigtig for stabiliseringen af 30 nm strukturen.
det endelige niveau af emballage er kendetegnet ved 700 nm-strukturen Seti metafasekromosomet. Det kondenserede stykke kromatin har enkarakteristisk stilladsstruktur, der kan detekteres i metafasekromosomer. Dette synes at være resultatet af omfattende looping af Dnai kromosomet.
de sidste definitioner, der skal præsenteres, er euchromatin ogheterochromatin., Når kromosomer er farvet med farvestoffer, ser de ud til at havevekslende let og mørkt farvede områder. De letfarvede regionerer euchromatin og indeholder enkeltkopi, genetisk aktivt DNA. De mørkt farvede områder er heterochromatin og indeholder gentagne sekvenser, der er genetisk inaktive.
centromerer og telomerer
centromerer og telomerer er to væsentlige træk ved alle eukaryote kromosomer. Hver giver en unik funktion, der er absolut nødvendig for kromosomets stabilitet., Centromerer er nødvendige for adskillelse af centromere under meiose og mitose, og teleomerer giver terminal stabilitet til kromosomet og sikrer dets overlevelse.
centromerer er de kondenserede regioner inden for kromosomet, somer ansvarlige for den nøjagtige adskillelse af det replikerede kromosomunder mitose og meiose. Når kromosomer er farvede, viser de typisk en mørkfarvet region, der er centromeren. Under mitose skal centromeren, der deles af søsterkromatiderne, opdele sig, så kromatiderne kan migrere tilmodsatte poler i cellen., På den anden side under den første meiotiske divisioncentromeren af søsterkromatider skal forblive intakt, mens de under meiosisII skal fungere som de gør under mitose. Derfor er centromeren en vigtig bestanddel af kromosomstruktur og adskillelse.
inden for centromere-regionen har de fleste arter flere steder, hvor spindelfibre fastgøres, og disse steder består af DNA såvel som protein. Den faktiske placering, hvor den vedhæftede fil forekommer kaldes kinetochore og er sammensat af både DNA og protein. DNA-sekvensen i disse regioner kaldescen DNA., Da CEN-DNA kan flyttes fra et kromosom til et andet og stadig give kromosomet evnen til at adskille, må disse sekvenser ikke give nogen anden funktion.
Typisk CEN DNA er omkring 120 basepar lang og består af flere sub-domæner, CVU-jeg, CVU-II og CVU-III. Mutationer i de første to sub-domæner har ingen effekt på adskillelse, men et punkt mutation i CVU-III sub-domæne, som helt eliminerer muligheden af centromer til at fungere i løbet af kromosom adskillelse., Derfor skal CDE-III være aktivt involveret i bindingen af spindelfibrene til centromeren.
proteinkomponenten i kinetochoren karakteriseres først nu. Et kompleks af tre proteiner kaldet Cbf-III binder til normale CDE-III-regioner, men kan ikke binde til en CDE-III-region med en punktmutation, der forhindrer mitotisk adskillelse. Desuden eliminerer mutanter af generne, der koder for CBF-III-proteinerne, også kromosomernes evne til at adskille sig under mitose., Yderligere analyser af DNA-og proteinkomponenterne i centromeren er nødvendige for fuldt ud at forstå mekanikken i kromosomsegregation.
telomerer er regionen af DNA i slutningen af det lineære eukaryotiskekromosom, der er nødvendige for replikation og stabilitet af kromosomet.McClintock anerkendte deres særlige træk, da hun bemærkede, at hvis tokromosomer blev brudt i en celle, kunne enden af den ene vedhæfte den andenog omvendt. Hvad hun aldrig observerede var fastgørelsen af den brudteende til slutningen af et ubrudt kromosom., Således enderne af brudte kromosomerer klæbrige, mens den normale ende ikke er klæbrig, hvilket tyder på enderne afkromosomer har unikke egenskaber. Normalt, men ikke altid, er telomericDNA heterokromatisk og indeholder direkte tandemly gentagne sekvenser. Den følgende tabel viser gentagne sekvenser af flere arter. Disse er ofte af formen (t / a) xGy hvor x er mellem 1 og 4 og y er større end 1.,
Telomere Repeat Sequences
| Species | Repeat Sequence |
|---|---|
| Arabidopsis | TTTAGGG |
| Human | TTAGGG |
| Oxytricha | TTTTGGGG |
| Slime Mold | TAGGG |
| Tetrahymena | TTGGGG |
| Trypanosome | TAGGG |
| Yeast | (TG)1-3TG2-3 |
Notice that the number of TG sequences and the number of cytosines in the yeast sequence varies., I det mindste for gær har det vist sig, at forskellige stammer indeholder forskellige længder af teleomerer, og at længden er under genetisk kontrol.
den primære vanskelighed med telomerer er replikationen af den laggende streng.Fordi DNA-syntese kræver en RNA-skabelon (der tilvejebringer den frie 3′-OHgroup) til primær DNA-replikation, og denne skabelon til sidst nedbrydes, ville en kort enkeltstrenget region blive efterladt i slutningen af kromosomet. Denne region ville være modtagelig for en .ymer, der nedbryder enkeltstrenget DNA., Resultatet ville være, at længden af kromosomet ville blive forkortet efter hver division. Men dette ses ikke.
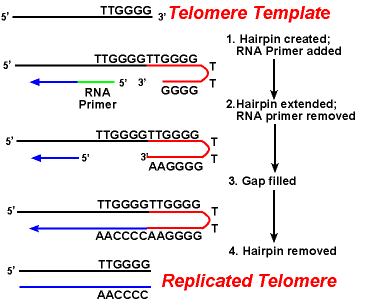
virkningen af telomeraseen .ymerne sikrer, at enderne af de hængende tråde replikeres korrekt. Et godt undersøgt system involverer Tetrahymena-proto .oerorganismen. Telomererne af denne organisme slutter i sekvensen 5 ‘-ttggg-3’. Telomerasen tilføjer en række 5′-ttggg-3′ – gentagelser til enderne af den lagrende streng. En hårnål opstår, når usædvanlige basepar mellem guaninrester i gentagelsesformen., Derefter fjernes RNA-primeren, og 5′ – enden af den hængende streng kan bruges til DNA-syntese. Ligation opstår mellem den færdige hængende streng og hårnålen. Endelig fjernes hårnålen ved 5′-ttggg-3 ‘ – gentagelsen. Således replikeres slutningen af kromosomet trofast. Følgende figur viser disse trin.,
Replikation af Telomerer
Analyse af DNA-Sekvenser i Eukaryote Genomer
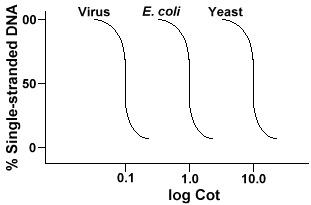
Den teknik, der er brugt til at bestemme rækkefølgen kompleksitet af alle genomeinvolves denaturering og renaturation af DNA. DNA denatureres ved opvarmning, der smelter H-bindingerne og gør DNA ‘ et enkeltstrenget. Hvisdnaet afkøles hurtigt, forbliver DNA ‘ et enkeltstrenget. Men hvis DNAis får lov til at køle langsomt, sekvenser, der er komplementære, finder hinanden og til sidst basepar igen., Den hastighed, hvormed DNA reanneals (en anden betegnelse for renature) er en funktion af de arter, hvorfra DNA .as isoleret. Nedenfor er en kurve, der opnås fra et simpelt genom.
Y-aksen er procentdelen af DNA ‘ et, der forbliver enkeltstrenget. Dette udtrykkes som et forhold mellem koncentrationen af enkeltstrenget DNA (C)og den totale koncentration af start-DNA ‘ et (Co). X-aksen isa log-skala af produktet af den indledende koncentration af DNA (i mol/liter)multipliceret med længden af tid reaktionen fortsatte (i sekunder)., Betegnelsenfor denne værdi er Cot og kaldes” Cot ” værdien. Selve kurvenkaldes en” Cot ” kurve. Som det kan ses kurven er temmelig glat whichhichindikates at reannealing sker langsommere, men gradvist over en periode oftime. En særlig værdi, der er nyttig, er Cot ,, dencot-værdi, hvor halvdelen af DNA ‘ et er reannealed.
trin involveret i DNA Denaturerings-og Renatureringsforsøg
1. Skær DNA ‘ et til en størrelse på omkring 400 bp.
2. Denaturere DNA ved opvarmning til 100oC.
3. Langsomt afkøles og tage prøver på forskellige tidsintervaller.
4., Bestem det % enkeltstrengede DNA på hvert tidspunkt.
form af en “Seng” kurve for en given art er en funktion af to faktorer:
- størrelsen eller kompleksiteten af genomet; og
- mængden af gentagne DNA i genomet
Hvis vi plotte “Cot” kurver af genomet af tre arter som bacteriophagelambda, E. coli og gær vil vi se, at de har samme form,men den Cot½ af gær vil være størst, E. coli nextand lambda mindste., Fysisk, jo større genomstørrelsen er, jo længere vil det tage for en sekvens at støde på dens komplementære sekvens i løsningen. Dette skyldes, at to komplementære sekvenser skal mødehinanden, før de kan parre. Jo mere komplekst genomet er, det vil sigemere unikke sekvenser, der er tilgængelige, jo længere tid vil det tage for nogento komplementære sekvenser at møde hinanden og parre. I betragtning af lignende koncentrationer i opløsning vil det tage en mere kompleks art længere at nå Cot ..,
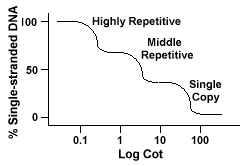
Gentagne DNA-sekvenser, DNA-sekvenser, der er fundet mere end én gang i afkodningen af de arter, har en markant indvirkning på “Cot” kurver.Hvis en bestemt sekvens er repræsenteret to gange i genomet, det vil have twocomplementary sekvenser til at parre med, og som sådan vil have en Barneseng værdi halfas stor som en sekvens, der er repræsenteret kun én gang i genomet.
eukaryote genomer har faktisk en bred vifte af sekvenser, der er repræsenteretpå forskellige niveauer af gentagelse., Enkeltkopisekvenser findes en gangeller et par gange i genomet. Mange af de sekvenser, der koder functionalgener falder ind i denne klasse. Midterste gentagne DNA findes fra 10s -1000 gange i genomet. Eksempler på disse ville omfatte rRNA og trnagener og opbevaringsproteiner i planter som majs. Midterste gentagne Dnakan variere fra 100-300 bp til 5000 bp og kan spredes gennem hele genomet.De mest rigelige sekvenser findes i den meget gentagne DNAclass., Disse sekvenser findes fra 100.000 til 1 million gange i genometog kan variere i størrelse fra et par til flere hundrede baser i længden. Disse sekvenser findes i områder af kromosomet,såsom heterochromatin, centromerer og telomerer og har tendens til at blive arrangeret som en tandem gentagelser. Denfølgende er et eksempel på en tandemly gentagne sekvens:
ATTATA ATTATA ATTATA // ATTATA
Genomer, der indeholder disse forskellige klasser af sekvenser reanneal i en differentmanner end genomer med kun en enkelt kopi sekvenser., I stedet for at have asingle glat” Cot ” kurve, tre forskellige kurver kan ses, hver repræsenterer en anden gentagelse klasse. De første sekvenser, for at reanneal er highlyrepetitive sekvenser, fordi så mange kopier af dem findes i genomet,og fordi de har en lav sekvens kompleksitet. Den anden del af genomet til reanneal er det midterste gentagne DNA, og den sidste del toreaneal er det enkelte kopi DNA., Følgende diagram viser “Cot”kurven for en “typisk” eukaryote genom
Den følgende tabel giver sekvensen fordelingen for udvalgte arter.
| Arter | Sekvens Distribution |
|---|---|
| Bakterier | 99.,27% Midten Gentagne 8% Meget Repetitive |
| Majs | 30% Enkelt Kopi 40% Midten Gentagne 20% Meget Repetitive |
| Hvede | 10% Enkelt Kopi 83% Midten Gentagne 4% Meget Repetitive |
| Arabidopsis | 55% Enkelt Kopi 27% Midten Gentagne 10% Meget Repetitive |
Sekvens Interspersion
Selv om genomer fra højere organismer indeholder en enkelt kopi, middlerepetitive og meget repeterende DNA-sekvenser, disse sekvenser er ikke arrangedsimilarly i alle arter., Det fremtrædende arrangement kaldes kort periodeinterspersion. Dette arrangement er kendetegnet ved gentagne sekvenser100-200 bp i længden spredt blandt enkeltkopisekvenser, der er 1000-2000bp i længden. Dette arrangement findes hos dyr, svampe og planter.
den anden type arrangement er langvarig interspersion. Dette er kendetegnet ved 5000 bp strækninger af gentagne sekvenser intersperedinithin regioner af 35.000 bp enkelt kopi DNA. Drosophila er et Eksempelaf en art med denne usædvanlige sekvens arrangement., I begge tilfælde ergentagne sekvenser normalt fra den midterste gentagne klasse. Vi diskuterede ovenfor, hvor der findes meget gentagne sekvenser.eukaryote Kromosomkaryotype
mens bakterier kun har et enkelt kromosom, har eukaryote arter mindst et par kromosomer. De fleste har mere end et par. Et andet relevant punkt er, at eukaryote kromosomer kun opdages under celledivision og ikke i alle stadier af cellecyklussen. De er i deres mestkondenserede form under metafase, når søsterkromatiderne er fastgjort.,Dette er det primære stadium, når cytogenetisk analyse udføres.
hver art er kendetegnet ved en karyotype. Karyotypen er enbeskrivelse af antallet af kromosomer i den normale diploide celle, såvel som deres størrelsesfordeling. For eksempel har det menneskelige kromosom 23 paraf kromosom, 22 somatiske par og et par kønskromosomer. Et vigtigt aspekt af genetisk forskning er at korrelere ændringer i karyotypen med ændringer i individets fænotype.
et vigtigt aspekt af genetik er korrelerende ændringer i karyotype medændringer i fænotype., For eksempel mennesker, der har et ekstra kromosom 21har do .ns syndrom. Insertioner, sletninger og ændringer i kromosomnummerkan detekteres af den dygtige cytogenetiker, men korrelerer disse medspecifikke fænotyper er vanskelige.
den første diskriminerende parameter ved udvikling af en karyotype er størrelsenog antallet af kromosomer. Selv om dette er nyttigt, giver det ikkenok detalje at være begynde udviklingen af en sammenhæng mellem struktur og funktion (fænotype)., For yderligere at skelne mellem kromosomer, debehandles med et farvestof, der pletter DNA ‘ et på en reproducerbar måde. Efterfarvning er nogle af regionerne let farvede, og andre er tungefarvede. Som beskrevet ovenfor kaldes de let farvede områderdeuchromatin, og den mørke farvede region kaldesheterochromatin. Det nuværende farvestof af chose er Giemsa-pletten, og det resulterende mønster kaldes G-banding-mønsteret.,
c-værdi paradoks
ud over at beskrive genomet af en organisme ved dets antal kromosomer,beskrives det også af mængden af DNA i en haploidcelle. Dette udtrykkes normalt som mængden af DNA pr. haploidcelle (normalt udtrykt som picogrammer)eller antallet af kilobaser pr.haploidcelle og kaldes C-værdien. Enumiddelbart træk ved eukaryote organismer fremhæver en specifik anomali, somblev påvist tidligt i molekylær forskning., Selvom eukaryote organismervises til at have 2-10 gange så mange gener som prokaryoter, de har mange orderaf størrelsesorden mere DNA i cellen. Genomer korreleret ikke med den formodede evolutionære kompleksitet af en art.Dette angives som C-værdien paradoks: mængden af DNA i haploidcellen af en organisme er ikke relateret til dens evolutionære kompleksitet. (Anotherimportant punkt at huske på er, at der ikke er nogen sammenhæng mellem antallet af kromosomer og den formodede evolutionære kompleksitet anorganism.,)
C Values of Organisms Used in Genetic Studies
| Species | Kilobases/haploid genome |
|---|---|
| E. coli |
4.5 x 103 |
| Human |
3.0 x 106 |
| Drosophila |
1.7 x 105 |
| Maize |
2.0 x 106 |
| Aribidopsis |
7.,0 x 104 |
Et dramatisk eksempel på den række af C-værdier kan ses i anlægget kingdomwhere Arabidopsis repræsenterer den lave ende, og lily (1,0 x 10^8 kb/haploidgenome) den høje ende af kompleksitet. Vægtmæssigt er dette 0,07 picogramsper haploid Arabidopsis genom og 100 picograms pr.haploid lilygenome.
genom-det komplette sæt kromosomer arvet fra en enkelt forælder; den komplette DNA-komponent af et individ; definitionen udelukker ofteorganeller