structura cromozomului eucariot
lungimea ADN-ului din nucleu este mult mai mare decât dimensiunea compartimentuluiîn care este conținut. Pentru a se încadra în acest compartiment, ADN-ul trebuie să fiecondensat într-un fel. Se exprimă gradul în care ADN-ul este condensatca raport de ambalare.raportul de ambalare – lungimea ADN-ului împărțită la lungimea în careeste ambalat
de exemplu, cel mai scurt cromozom uman conține 4,6 x 107 bp de ADN(aproximativ 10 ori dimensiunea genomului E. coli)., Aceasta este echivalentă cu 14.000 µm de ADN extins. În starea sa cea mai condensată în timpul mitozei, cromozomul are o lungime de aproximativ 2 µm. Acest lucru oferă un raport de ambalare de 7000 (14.000 / 2).
pentru a obține raportul general de ambalare, ADN-ul nu este ambalat direct în structura finală a cromatinei. În schimb, conține mai multe ierarhii de organizare.Primul nivel de ambalare se realizează prin înfășurarea ADN-ului în jurul unui proteincore pentru a produce o structură „asemănătoare cu mărgele” numită nucleozom. Acestaoferă un raport de ambalare de aproximativ 6., Această structură este invariantă în ambeleeuchromatină și heterochromatină a tuturor cromozomilor. Cel de-al doilea nivel de packingis forma de margele într-o structură elicoidală numită de 30 nm fiberthat se găsește atât în interfază cromatinei și mitotice cromozomii. Thisstructure crește raportul de ambalare la aproximativ 40. Ambalarea finală occurswhen fibra este organizat în bucle, schele și domenii care dau un finalpacking raport de aproximativ 1000 în interfază cromozomii și aproximativ 10.000 de inmitotic cromozomi.,cromozomii Eucarioți constau dintr-un complex ADN-proteină care este organizat într-o manieră compactă care permite stocarea unei cantități mari de ADN în nucleul celulei. Desemnarea subunității cromozomului este cromatina.Unitatea fundamentală a cromatinei este nucleozomul.,romatin – unitatea de analiză a cromozomilor; cromatinei reflectsthe structura generală a unui cromozom, dar nu este unic pentru orice particularchromosome
Nucleosome – mai simple de ambalare structura ADN-ului care se găsește întoate eucariote cromozomii; ADN-ul este înfășurat în jurul unui octamer de mici basicproteins numite histone; 146 bp este înfășurat în jurul miezului și remainingbases link-ul de la următoarea nucleosome; această structură determină negative supercoiling
nucleosome este format din aproximativ 200 bp înfășurat în jurul unui histone octamerthat conține două copii de proteine histone H2A, H2B, H3 și H4., Acestea sunt cunoscute ca histone de bază. Histonele sunt proteine de bază care au o afinitatepentru ADN și sunt cele mai abundente proteine asociate cu ADN-ul. Secvența aminoacidă a acestor patru histone este conservată sugerând o funcție similarăpentru toți.
lungimea ADN-ului care este asociată cu unitatea nucleozomică variază întrespecii. Dar, indiferent de dimensiune, sunt implicate două componente ADN. CoreDNA este ADN-ul care este de fapt asociat cu histone octamer.Această valoare este invariantă și este de 146 perechi de baze., Miezul ADN-ul formează două loopsaround la octamer, și acest lucru permite două regiuni care sunt 80 bp în afară tobe adus în imediata apropiere. Astfel, două secvențe care sunt îndepărtate potinteracționează cu aceeași proteină de reglementare pentru a controla expresia genelor. Adna care se află între fiecare octamer histone se numește ADN-ul Linker și poate varia în lungime de la 8 la 114 perechi de baze. Această variație este specifică, dar variația lungimii ADN-ului linker a fost, de asemenea, asociată cu stadiul de dezvoltare al organismului sau regiuni specifice ale genomului.,
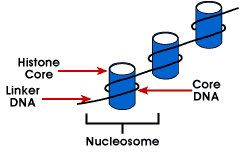
La următorul nivel de organizare al cromatinei este fibra de 30 nm. Thisappears a fi o structură solenoid cu aproximativ 6 nucleozomi pe rândul său. Thisgives o ambalare raport de 40, ceea ce înseamnă că la fiecare 1 µm de-a lungul axiscontains 40 µm a ADN-ului. Stabilitatea acestei structuri necesităprezența ultimului membru al familiei genei histone, histone H1., Becauseexperiments striptease H1 din cromatinei menține nucleosome, dar nu 30 nm structura, s-a concluzionat că H1 este important pentru stabilizationof 30 nm structura.nivelul final al ambalajului este caracterizat de structura de 700 nm văzutăîn cromozomul metafazei. Piesa condensată de cromatină are o structură de schele caracteristică care poate fi detectată în metafasechromozomi. Acest lucru pare a fi rezultatul unei bucle extinse a Dnuluiîn cromozom.ultimele definiții care trebuie prezentate sunt euchromatină șiheterocromatină., Când cromozomii sunt colorați cu coloranți, par să aibăalternând regiuni ușor și întunecate. Regiunile ușor pătate sunt euchromatină și conțin ADN-ul unic, activ genetic. Regiunile întunecate sunt heterocromatine și conțin secvențe repetitivecare sunt inactive genetic.Centromerii și telomerii Centromerii și telomerii sunt două caracteristici esențiale ale tuturor cromozomilor eucarioți. Fiecare oferă o funcție unică care este absolut necesară pentru stabilitatea cromozomului., Centromerii sunt necesari pentru segregarea centromerului în timpul meiozei și mitozei, iar teleomerii asigură stabilitatea terminală a cromozomului și asigură supraviețuirea acestuia.centromerele sunt acele regiuni condensate din cromozomul caresunt responsabile pentru segregarea exactă a cromozomului replicatîn timpul mitozei și meiozei. Când cromozomii sunt colorați, aceștia prezintă de obicei o regiune întunecată, care este centromerul. În timpul mitozei, centromerul care este împărțit de cromatidele surori trebuie să se împartă astfel încât cromatidele să poată migra la poli opuși ai celulei., Pe de altă parte, în prima meiotice divisionthe centromerului sora cromatide trebuie să rămână intact, întrucât în timpul meiosisII ei trebuie să acționeze așa cum au făcut în timpul mitozei. Prin urmare, centromerul este o componentă importantă a structurii cromozomilor și a segregării.în cadrul regiunii centromere, majoritatea speciilor au mai multe locații în care se atașează fibrele axului, iar aceste site-uri constau atât din ADN, cât și din proteine. Locația reală în care are loc atașamentul se numește kinetochore și este compusă atât din ADN, cât și din proteine. Secvența ADN din aceste regiuni este numităcen ADN., Deoarece ADN-ul CEN poate fi mutat de la un cromozom la altul și oferă în continuare cromozomului capacitatea de a segrega, aceste secvențe nu trebuie să ofere nicio altă funcție.de obicei, ADN-ul CEN are o lungime de aproximativ 120 de perechi de baze și constă din mai multe subdomenii, CDE-i, CDE-II și CDE-III. mutațiile din primele două subdomenii nu au efect asupra segregării, dar o mutație punctuală în subdomeniul CDE-III elimină complet capacitatea centromerului de a funcționa în timpul segregării cromozomilor., Prin urmare, CDE-III trebuie să fie implicat activ în legarea fibrelor axului la centromer.componenta proteică a kinetochorului este caracterizată abia acum. Un complex de trei proteine numite Cbf-III se leagă de regiunile normale CDE-III, dar nu se poate lega de o regiune CDE-III cu o mutație punctuală care împiedică segregarea mitotică. Mai mult, mutanții genelor care codifică proteinele Cbf-III elimină, de asemenea, capacitatea cromozomilor de a se separa în timpul mitozei., Analizele suplimentare ale componentelor ADN și proteice ale centromerului sunt necesare pentru a înțelege pe deplin mecanica segregării cromozomilor.telomerii sunt regiunea ADN – ului la sfârșitul eucarioticăcromozomul liniar care este necesar pentru replicarea și stabilitatea cromozomului.McClintock și-a recunoscut trăsăturile Speciale când a observat, că dacă douăochromozomii au fost rupți într-o celulă, sfârșitul unuia s-ar putea atașa la celălaltși invers. Ceea ce nu a observat niciodată a fost atașarea ruptuluipână la sfârșitul unui cromozom neîntrerupt., Astfel, capetele cromozomilor rupți sunt lipicioase, în timp ce capătul normal nu este lipicios, sugerând că capetele cromozomilor au caracteristici unice. De obicei, dar nu întotdeauna, telomericDNA este heterocromatică și conține secvențe directe tandemly repetate. Tabelul următor prezintă secvențele repetate ale mai multor specii. Acestea sunt adesea de forma (T/A)xGy unde x este între 1 și 4 și y este mai mare decât 1.,
Telomere Repeat Sequences
| Species | Repeat Sequence |
|---|---|
| Arabidopsis | TTTAGGG |
| Human | TTAGGG |
| Oxytricha | TTTTGGGG |
| Slime Mold | TAGGG |
| Tetrahymena | TTGGGG |
| Trypanosome | TAGGG |
| Yeast | (TG)1-3TG2-3 |
Notice that the number of TG sequences and the number of cytosines in the yeast sequence varies., Cel puțin pentru drojdie, s-a demonstrat că diferite tulpini conțin lungimi diferite de teleomere și că lungimea este sub control genetic.
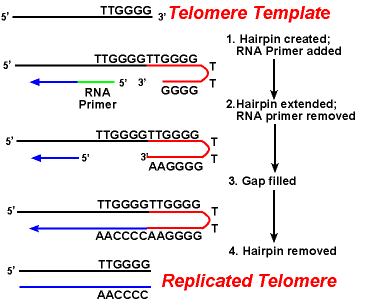
dificultatea principală cu telomerii este replicarea firului rămas.Deoarece sinteza ADN-ului necesită un șablon ARN (care oferă gratuit 3′-OHgroup) pentru replicarea ADN-ului prim, iar acest șablon este în cele din urmă degradat, o regiune scurtă monocatenară ar fi lăsată la sfârșitul cromozomului. Această regiune ar fi susceptibilă la enzime care degradează ADN-ul monocatenar., Rezultatul ar fi că lungimea cromozomului ar fi scurtată după fiecare diviziune. Dar acest lucru nu este văzut.acțiunea enzimelor telomerazei asigură că capetele firelor rămase sunt reproduse corect. Un sistem bine studiat implică organismul protozoare Tetrahymena. Telomerii acestui organism se termină în secvența 5′ – TTGGGG-3′. Telomeraza adaugă o serie de 5′-TTGGGG-3′ repetă la capetele firului rămas. Un ac de păr apare atunci când perechi neobișnuite de bază între reziduurile de guanină în forma repetată., Apoi grundul ARN este îndepărtat, iar capătul 5′ al firului rămas poate fi utilizat pentru sinteza ADN-ului. Ligarea are loc între firul final rămas și acul. În cele din urmă, acul este îndepărtat la repetarea 5′-TTGGGG-3′. Astfel, sfârșitul cromozomului este replicat fidel. Următoarea figură prezintă acești pași.,
Replicarea Telomerelor
Analiză de Secvențe ADN la Eucariote Genomul
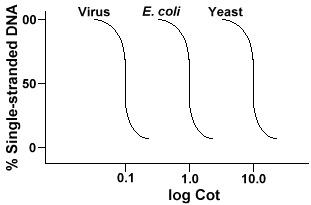
tehnica care este folosit pentru a determina secvența de orice complexitate genomeinvolves la denaturarea și renaturarea ADN-ului. ADN-ul este denaturat prin încălzire care topește legăturile H și face ADN-ul monocatenar. Dacă ADN-ul este răcit rapid, ADN-ul rămâne monocatenar. Dar dacă ADN-ul este lăsat să se răcească încet, secvențele care sunt complementare se vor găsi reciproc și, în cele din urmă, se vor împerechea din nou., Rata la care ADN-ul reanneals(un alt termen pentru renature) este o funcție de specia de la care DNAwas izolate. Mai jos este o curbă care este obținută dintr-un genom simplu.
axa Y este procentul de ADN care rămâne singur catenar. Aceasta esteexprimată ca raport între concentrația ADN-ului monocatenar (C) și concentrația totală a ADN-ului de pornire (Co). Axa X esteo scară log a produsului concentrației inițiale de ADN (în moli/litru)înmulțită cu durata de timp în care reacția a continuat (în secunde)., Desemnarea pentru această valoare este pătuț și se numește valoarea „pătuț”. Curba în sinese numește curbă „pat”. După cum se poate observa curba este destul de netedă, ceea ce indică faptul că reanalizarea are loc încetinind, dar treptat, pe o perioadă de timp. O valoare specială care este utilă este Cot½, valoarea pot în cazul în care jumătate din ADN-ul a reanalizat.etapele implicate în experimentele de denaturare și renaturare a ADN-ului
1. Tăiați ADN-ul la o dimensiune de aproximativ 400 bp.
2. Denatura ADN-ul prin încălzire la 100oC.
3. Se răcește încet și se prelevează probe la intervale diferite de timp.
4., Determinați ADN-ul monocatenar % la fiecare moment.
forma un „Pat” curba pentru o anumită specie este o funcție de doi factori:
- dimensiunea sau complexitatea genomului; și
- cantitatea de ADN repetitiv în genom
Dacă ne complot „Pat” curbe de genomul trei specii, cum ar fi bacteriophagelambda, E. coli și drojdie, vom vedea că acestea au aceeași formă,dar Cot½ de drojdie va fi mai mare, E. coli next lambda mai mic., Fizic, cu cât dimensiunea genomului este mai mare, cu atât va dura mai mult pentru ca o secvență să întâlnească secvența complementară în soluție. Acest lucru se datorează faptului că două secvențe complementare trebuie să se întâlneascăunul pe altul înainte de a se putea asocia. Mai complexe genomului, care este de mai multe secvențe unice, care sunt disponibile, cu atât mai mult va dura pentru anytwo secvențe complementare de a întâlni reciproc și pereche. Dat similarconcentrations în soluție, se va lua apoi o specie mult mai complexă longerto ajunge Cot½ .,
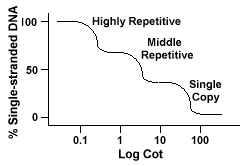
secvențele de ADN Repetate, secvente de ADN care sunt găsite mai mult de o dată în genomul speciei, au distinctiv efecte pe „Pat” curbe.Dacă o secvență specifică este reprezentată de două ori în genomul va avea twocomplementary secvențe pentru a asocia cu și ca atare va avea un Pat valoare atat de mare ca o secvență a reprezentat doar o dată în genom.genomii Eucarioți au de fapt o gamă largă de secvențe care sunt reprezentatela diferite niveluri de repetare., Secvențele cu o singură copie se găsesc o datăsau de câteva ori în genom. Multe dintre secvențele care codifică funcționalgene se încadrează în această clasă. ADN-ul repetitiv Mijlociu se găsește de la 10s -1000 ori în genom. Exemple de acestea ar include ARNr și trnagene și proteine de stocare în plante, cum ar fi porumbul. Mediu repetitiv DNAcan variază de la 100-300 bp la 5000 bp și poate fi dispersat în întregul genom.Secvențele cele mai abundente se găsesc în Dnaclasa extrem de repetitivă., Aceste secvențe se găsesc de la 100.000 la 1 milion de ori în genomși poate varia în mărime de la câteva până la câteva sute de baze în lungime. Aceste consecințe se găsesc în regiuni ale cromozomului,cum ar fi heterochromatina, centromerii și telomerii și tind să fie aranjate ca repetiții tandem. Următoarele este un exemplu de un tandemly repetă secvența:
ATTATA ATTATA ATTATA // ATTATA
Genomul care conțin aceste diferite clase de secvențe reanneal într-un differentmanner decât genomul cu doar un singur exemplar secvențe., În loc de a avea o singură curbă netedă „pat”, trei curbe distincte pot fi văzute, fiecare reprezentând o clasă de repetiție diferită. Primele secvențe care se reanalizează sunt secvențele highlyrepetitive, deoarece există atât de multe copii ale acestora în genom și pentru că au o complexitate redusă a secvențelor. A doua porțiune a genomului reanneal este ADN-ul repetitiv de mijloc, iar porțiunea finală toreanneal este ADN-ul cu o singură copie., Următoarea diagramă descrie „Pat”curba pentru un „tipic” eucariote genom
tabelul următor prezintă secvența de distribuție pentru speciile selectate.
| specii | distribuție secvență |
|---|---|
| bacterii | 99.,27% din Mijloc Repetitive 8% Extrem de Repetitive |
| Porumb | 30% Singur Exemplar 40% Mijlocul Repetitive 20% Extrem de Repetitive |
| de Grâu | 10% Singur Exemplar 83% Mijlocul Repetitive 4% Extrem de Repetitive |
| Arabidopsis | 55% Singur Exemplar 27% din Mijloc Repetitive 10% Extrem de Repetitive |
Secvență Interspersion
Chiar dacă genomurile organismelor superioare conțin singur exemplar, middlerepetitive și extrem de secvențe repetitive de ADN, aceste secvențe nu sunt arrangedsimilarly la toate speciile., Aranjamentul proeminent se numește perioadă scurtăinterspersiune. Acest aranjament este caracterizat prin secvențe repetate100-200 BP în lungime intercalate între secvențe de o singură copie, care sunt 1000-2000bp în lungime. Acest aranjament se găsește la animale, ciuperci și plante.
al doilea tip de aranjament este intercalarea pe termen lung. Aceasta se caracterizează prin întinderi de 5000 bp de secvențe repetate intercalate în regiuni de 35.000 bp de ADN cu o singură copie. Drosophila este un exemplua unei specii cu acest aranjament de secvență neobișnuit., În ambele cazuri, secvențele repetate sunt de obicei din clasa repetitivă de mijloc. Am discutat mai sus unde se găsesc secvențe foarte repetitive.în timp ce bacteriile au doar un singur cromozom, speciile eucariote au cel puțin o pereche de cromozomi. Majoritatea au mai mult de o pereche. Un alt relevantpunct este că cromozomi eucariote sunt detectate apar numai în timpul celldivision și nu în timpul tuturor etapelor ciclului celular. Ele sunt în cele mai multeforma condensată în timpul metafazei când sunt atașate cromatidele surori.,Aceasta este etapa primară când se efectuează analiza citogenetică.fiecare specie este caracterizată de un cariotip. Cariotipul este adescrierea numărului de cromozomi din celula diploidă normală, precum șica distribuția dimensiunii lor. De exemplu, cromozomul uman are 23 de perechide cromozom, 22 de perechi somatice și o pereche de cromozomi sexuali. Un aspect important al cercetării genetice este corelarea modificărilor cariotipului cu schimbărileîn fenotipul individului.un aspect important al geneticii este corelarea modificărilor cariotipuluimodificări ale fenotipului., De exemplu, oamenii care au un cromozom suplimentar 21au sindromul Down. Inserțiile, ștergerile și modificările numărului cromozomilor pot fi detectate de către citogeneticianul calificat, dar corelarea acestora cu fenotipurile specifice este dificilă.primul parametru discriminatoriu la dezvoltarea unui cariotip este dimensiuneași numărul cromozomilor. Deși acest lucru este util, nu oferă suficiente detalii pentru a începe dezvoltarea unei corelații între structură și funcție (fenotip)., Pentru a distinge în continuare între cromozomi, eisunt tratate cu un colorant care pătează ADN-ul într-o manieră reproductibilă. După colorare, unele regiuni sunt ușor colorate, iar altele sunt greu colorate. După cum este descris mai sus, puțin pătat regiuni sunt calledeuchromatin, și întuneric colorate regiune este calledheterochromatin. Colorantul actual al ales este pata Giemsa, iar modelul rezultat se numește modelul G-banding.,pe lângă descrierea genomului unui organism prin numărul său de cromozomi,este descris și de cantitatea de ADN dintr-o celulă haploidă. Aceasta este de obiceiexprimată ca cantitatea de ADN pe celulă haploidă (de obicei exprimată ca picograme)sau numărul de kilobaze pe celulă haploidă și se numește valoarea C. O caracteristică imediată a organismelor eucariote evidențiază o anomalie specificăa fost detectată la începutul cercetării moleculare., Chiar dacă organismele eucariote par să aibă de 2-10 ori mai multe gene decât procariote, ele au multe ordine de mărime mai mult ADN în celulă. Mai mult, cantitatea de ADN pe genomnu este corelată cu presupusa complexitate evolutivă a unei specii.Acest lucru este declarat ca paradoxul valorii C: Cantitatea de ADN din haploidcell a unui organism nu este legată de complexitatea sa evolutivă. (Un alt punct important de reținut este că nu există nicio relație între numărul de cromozomi și presupusa complexitate evolutivă a anorganismului.,)
C Values of Organisms Used in Genetic Studies
| Species | Kilobases/haploid genome |
|---|---|
| E. coli |
4.5 x 103 |
| Human |
3.0 x 106 |
| Drosophila |
1.7 x 105 |
| Maize |
2.0 x 106 |
| Aribidopsis |
7.,0 x 104 |
Un exemplu dramatic de gama de C valorile pot fi văzute în plantă kingdomwhere Arabidopsis reprezintă low-end și lily (1,0 x 10^8 kb/haploidgenome) sfârșitul ridicat de complexitate. In termenii de greutate este 0.07 picogramsper haploid genomului la Arabidopsis și 100 picograme per haploid lilygenome.genomul-setul complet de cromozomi moșteniți de la un singur părinte;componenta ADN completă a unui individ; definiția exclude adeseaorganele